Eiwitfosforylering regelt de meeste cellulaire processen, inclusief maar niet beperkt tot de reactie op DNA-schade, groeifactorsignalering en de passage door mitose 1,2,3. In zoogdiercellen worden de meeste eiwitten op een bepaald moment gefosforyleerd op een of meer serine-, threonine- of tyrosineresiduen, waarbij fosfostropenen en fosfoothreonines ongeveer 98% van alle fosforyleringsplaatsen uitmaken 2,3. Hoewel kinasen uitgebreid zijn bestudeerd in cellulaire signalering, is de rol van PPP’s in de regulatie van dynamische cellulaire processen nog steeds in opkomst.
Fosforyleringsdynamiek wordt gecontroleerd door het dynamische samenspel tussen kinasen en fosfatasen. In zoogdiercellen zijn er meer dan 400 eiwitkinasen die serine / threoninefosforylering katalyseren. Meer dan 90% van deze sites wordt gedefosforyleerd door fosfoproteïnefosfatasen (PPP’s), een kleine familie van enzymen die bestaat uit PP1, PP2A, PP2B, PP4-7, PPT en PPZ 2,3. PP1 en PP2A zijn verantwoordelijk voor de meerderheid van fosfosforrine en fosfotheonine defosforylering in een cel 2,3,4. Het opmerkelijke verschil in aantal tussen kinasen en fosfatasen en het gebrek aan specificiteit van PPP katalytische subeenheden in vitro leidden tot de overtuiging dat kinasen de belangrijkste determinant zijn van fosforylering 2,3. Meerdere studies hebben echter aangetoond dat fosfatasen substraatspecificiteit vaststellen door de vorming van multimere holo-enzymen 5,6,7,8,9. PP1 is bijvoorbeeld een heterodimeer dat bestaat uit een katalytische subeenheid en op een gegeven moment een van de meer dan 150 regulerende subeenheden 6,7,8. Omgekeerd is PP2A een heterotrimeer dat wordt gevormd uit een steiger (A), een regulerende (B) en een katalytische (C) subeenheid 2,3,9. Er zijn vier verschillende families van PP2A regulerende subeenheden (B55, B56, PR72 en striatine), elk met meerdere genen, splicevarianten en lokalisatiepatronen 2,3,9. Het multimere karakter van PPP’s vult de leemte in het aantal kinasen en PPP-katalytische subeenheden. Het creëert echter analytische uitdagingen voor het bestuderen van PPP-signalering. Om PPP-signalering uitgebreid te analyseren, is het van cruciaal belang om de verschillende holo-enzymen in een cel of weefsel te onderzoeken. Er is grote vooruitgang geboekt bij het bestuderen van het menselijke kinoom door het gebruik van kinaseremmerparels, multiplexremmerparels of kinobeads genoemd, een chemische proteomische strategie waarbij kinaseremmers worden geïmmobiliseerd op kralen en massaspectrometrie wordt gebruikt om verrijkte kinasen en hun interactoren te identificeren 10,11,12,13.
We hebben een vergelijkbare aanpak vastgesteld om PPP-biologie te bestuderen. Deze techniek omvat affiniteitsvangst van PPP-katalytische subeenheden met behulp van kralen met een geïmmobiliseerde, niet-selectieve PPP-remmer genaamd microcystine-LR (MCLR) genaamd fosfataseremmerparels (PIBs)14,15. In tegenstelling tot andere methoden die de endogene tagging of expressie van exogene PPP-subeenheden vereisen die de eiwitactiviteit of lokalisatie kunnen veranderen, maakt PIB-MS de verrijking mogelijk van endogene PPP-katalytische subeenheden, hun bijbehorende regulerende en steigerende subeenheden en interagerende eiwitten (het PPPome genoemd) uit cellen en weefsels op een bepaald tijdstip of onder specifieke behandelingsomstandigheden. MCLR remt PP1, PP2A, PP4-6, PPT en PPZ bij nanomolaire concentraties, waardoor PIB’s zeer effectief zijn in het verrijken voor de PPPome16. Deze methode kan worden geschaald voor gebruik op elk uitgangsmateriaal, van cellen tot klinische monsters. Hier beschrijven we in detail het gebruik van PIB’s en massaspectrometrie (PIB-MS) om het endogene PPPome en de modificatietoestanden efficiënt vast te leggen, te identificeren en te kwantificeren.
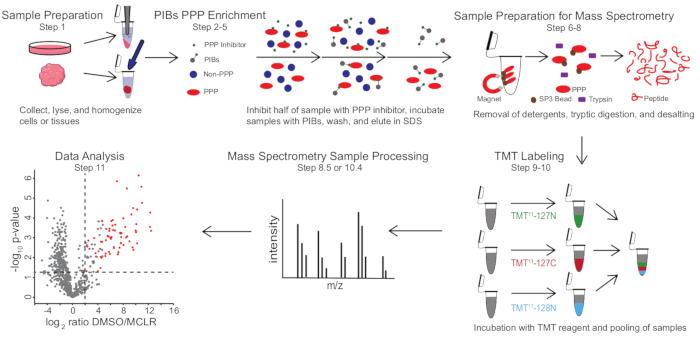
Figuur 1: Visuele samenvatting van het PIB-MS protocol. In een PIB-MS-experiment kunnen monsters worden verkregen in verschillende vormen, van cellen tot tumoren. Het monster wordt verzameld, gelyseerd en gehomogeniseerd voorafgaand aan PPP-verrijking. Ter verrijking voor PPP’s wordt het lysaat geïncubeerd met PIB’s met of zonder PPP-remmer, zoals MCLR. De PIB’s worden vervolgens gewassen en PPP’s worden geëlueerd in denatureringsomstandigheden. De monsters worden voorbereid voor massaspectrometrie-analyse door de verwijdering van detergentia door sp3-eiwitverrijking, tryptische vertering en ontzouting. Monsters kunnen dan optioneel TMT-gelabeld worden voorafgaand aan massaspectrometrie analyse. Klik hier om een grotere versie van deze figuur te bekijken.
PIB-MS omvat lysis en klaring van cellen of weefsels, incubatie van het lysaat met PIBs, elutie en analyse van het eluaat via western blotting of op massaspectrometrie gebaseerde benaderingen (figuur 1). De toevoeging van vrije MCLR kan worden gebruikt als een controle om specifieke PIB-bindmiddelen te onderscheiden van niet-specifieke interactoren. Voor de meeste toepassingen kan een labelvrije aanpak worden gebruikt om eiwitten in eluaten direct te identificeren. In gevallen waarin een grotere precisie in kwantificering of de identificatie van soorten met een lage abundantie nodig is, kan verdere verwerking met tandem mass-tag (TMT) -etikettering worden gebruikt om de dekking te vergroten en de input te verminderen.
