La phosphorylation des protéines contrôle la plupart des processus cellulaires, y compris, mais sans s’y limiter, la réponse aux dommages à l’ADN, la signalisation du facteur de croissance et le passage par la mitose 1,2,3. Dans les cellules de mammifères, la majorité des protéines sont phosphorylées à un ou plusieurs résidus de sérine, de thréonine ou de tyrosine à un moment donné, les phosphosérines et les phosphothréonines représentant environ 98 % de tous les sites de phosphorylation 2,3. Alors que les kinases ont été largement étudiées dans la signalisation cellulaire, le rôle des PPP dans la régulation des processus cellulaires dynamiques est encore émergent.
La dynamique de phosphorylation est contrôlée par l’interaction dynamique entre les kinases et les phosphatases. Dans les cellules de mammifères, il existe plus de 400 protéines kinases qui catalysent la phosphorylation de la sérine/thréonine. Plus de 90% de ces sites sont déphosphorylés par les phosphoprotéines phosphatases (PPP), une petite famille d’enzymes composée de PP1, PP2A, PP2B, PP4-7, PPT et PPZ 2,3. PP1 et PP2A sont responsables de la majorité de la déphosphosérine et de la phosphothréonine dans une cellule 2,3,4. La différence notable de nombre entre les kinases et les phosphatases et le manque de spécificité des sous-unités catalytiques PPP in vitro ont conduit à la croyance que les kinases sont le principal déterminant de la phosphorylation 2,3. Cependant, de multiples études ont montré que les phosphatases établissaient la spécificité du substrat par la formation d’holoenzymes multimériques 5,6,7,8,9. Par exemple, PP1 est un hétérodimère constitué d’une sous-unité catalytique et, à un moment donné, d’une des plus de 150 sous-unités régulatrices 6,7,8. Inversement, PP2A est un hétérotrimère formé d’un échafaudage (A), d’une sous-unité régulatrice (B) et d’une sous-unité catalytique (C) 2,3,9. Il existe quatre familles distinctes de sous-unités régulatrices PP2A (B55, B56, PR72 et striatine), chacune avec plusieurs gènes, variantes d’épissure et modèles de localisation 2,3,9. La nature multimérique des PPP comble le manque de kinases et de sous-unités catalytiques PPP. Cependant, cela crée des défis analytiques pour l’étude de la signalisation PPP. Pour analyser de manière exhaustive la signalisation PPP, il est essentiel d’étudier les différentes holoenzymes au sein d’une cellule ou d’un tissu. De grands progrès ont été réalisés dans l’étude du kinome humain grâce à l’utilisation de perles inhibitrices de kinases, appelées perles inhibitrices multiplex ou kinobeads, une stratégie protéomique chimique où les inhibiteurs de kinases sont immobilisés sur des perles et où la spectrométrie de masse est utilisée pour identifier les kinases enrichies et leurs interacteurs 10,11,12,13.
Nous avons établi une approche similaire pour étudier la biologie des PPP. Cette technique implique la capture d’affinité de sous-unités catalytiques PPP à l’aide de billes avec un inhibiteur de PPP non sélectif immobilisé appelé microcystine-LR (MCLR) appelé billes inhibitrices de phosphatase (PIBs)14,15. Contrairement à d’autres méthodes qui nécessitent le marquage endogène ou l’expression de sous-unités PPP exogènes susceptibles de modifier l’activité ou la localisation des protéines, LE PIB-MS permet l’enrichissement des sous-unités catalytiques PPP endogènes, de leurs sous-unités régulatrices et d’échafaudage associées et des protéines en interaction (appelées PPPome) à partir de cellules et de tissus à un moment donné ou dans des conditions de traitement spécifiques. McLR inhibe PP1, PP2A, PP4-6, PPT et PPZ à des concentrations nanomolaires, ce qui rend les PIB très efficaces pour enrichir le PPPome16. Cette méthode peut être mise à l’échelle pour être utilisée sur n’importe quel matériau de départ, des cellules aux échantillons cliniques. Ici, nous décrivons en détail l’utilisation des PIB et de la spectrométrie de masse (PIB-MS) pour capturer, identifier et quantifier efficacement le PPPome endogène et ses états de modification.
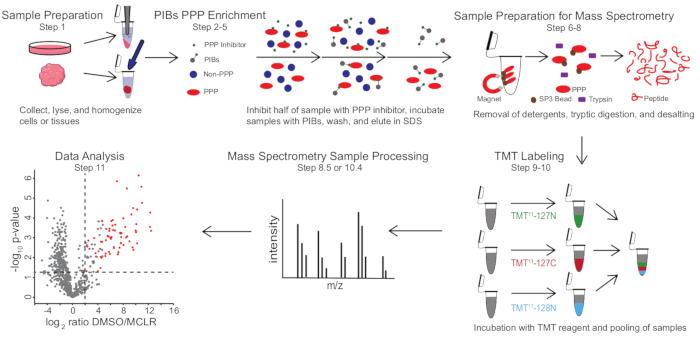
Figure 1 : Résumé visuel du protocole PIB-MS. Dans une expérience PIB-MS, des échantillons peuvent être obtenus sous diverses formes, des cellules aux tumeurs. L’échantillon est collecté, lysé et homogénéisé avant l’enrichissement en PPP. Pour enrichir les PPP, le lysat est incubé avec des PIB avec ou sans inhibiteur de PPP, tel que LE MCLR. Les PIB sont ensuite lavés et les PPP sont élués dans des conditions de dénaturation. Les échantillons sont préparés pour l’analyse par spectrométrie de masse par élimination des détergents par enrichissement en protéines SP3, digestion tryptique et dessalement. Les échantillons peuvent ensuite être éventuellement marqués TMT avant l’analyse par spectrométrie de masse. Veuillez cliquer ici pour voir une version agrandie de cette figure.
PIB-MS implique la lyse et la clarification des cellules ou des tissus, l’incubation du lysat avec des PIB, l’élution et l’analyse de l’éluat via des approches basées sur le transfert occidental ou la spectrométrie de masse (Figure 1). L’ajout de MCLR libre peut être utilisé comme contrôle pour distinguer des liants PIB spécifiques des interacteurs non spécifiques. Pour la plupart des applications, une approche sans étiquette peut être utilisée pour identifier directement les protéines dans les éluats. Dans les cas où une plus grande précision dans la quantification ou l’identification des espèces de faible abondance est nécessaire, un traitement supplémentaire avec l’étiquetage en tandem masse-étiquette (TMT) peut être utilisé pour augmenter la couverture et diminuer les intrants.
