Die Proteinphosphorylierung steuert die meisten zellulären Prozesse, einschließlich, aber nicht beschränkt auf die Reaktion auf DNA-Schäden, die Signalisierung des Wachstumsfaktors und die Passage durch Mitose 1,2,3. In Säugetierzellen wird die Mehrheit der Proteine zu einem bestimmten Zeitpunkt an einem oder mehreren Serin-, Threonin- oder Tyrosinresten phosphoryliert, wobei Phosphosrine und Phosphothreonine etwa 98% aller Phosphorylierungsstellen ausmachen 2,3. Während Kinasen in der zellulären Signalgebung ausführlich untersucht wurden, zeichnet sich die Rolle von PPPs bei der Regulation dynamischer zellulärer Prozesse noch ab.
Die Phosphorylierungsdynamik wird durch das dynamische Zusammenspiel von Kinasen und Phosphatasen gesteuert. In Säugetierzellen gibt es mehr als 400 Proteinkinasen, die die Serin/Threonin-Phosphorylierung katalysieren. Über 90% dieser Stellen werden durch Phosphoproteinphosphatasen (PPPs) dephosphoryliert, eine kleine Familie von Enzymen, die aus PP1, PP2A, PP2B, PP4-7, PPT und PPZ 2,3 besteht. PP1 und PP2A sind für den Großteil der Phosphoserin- und Phosphothreonin-Dephosphorylierung innerhalb einer Zelle verantwortlich 2,3,4. Der bemerkenswerte Unterschied in der Anzahl zwischen Kinasen und Phosphatasen und die mangelnde Spezifität der PPP-katalytischen Untereinheiten in vitro führten zu der Annahme, dass Kinasen die Hauptdeterminante der Phosphorylierungsind 2,3. Mehrere Studien haben jedoch gezeigt, dass Phosphatasen die Substratspezifität durch die Bildung multimerer Holoenzyme 5,6,7,8,9 etablieren. Zum Beispiel ist PP1 ein Heterodimer, das aus einer katalytischen Untereinheit und zu einem bestimmten Zeitpunkt aus einer der mehr als 150 regulatorischen Untereinheiten 6,7,8 besteht. Umgekehrt ist PP2A ein Heterotrimer, der aus einem Gerüst (A), einer regulatorischen (B) und einer katalytischen (C)Untereinheit 2,3,9 besteht. Es gibt vier verschiedene Familien von regulatorischen PP2A-Untereinheiten (B55, B56, PR72 und Striatin), jede mit mehreren Genen, Spleißvarianten und Lokalisationsmustern 2,3,9. Die multimere Natur von PPPs füllt die Lücke in der Anzahl der Kinasen und PPP-katalytischen Untereinheiten. Es schafft jedoch analytische Herausforderungen für die Untersuchung der PPP-Signalgebung. Um die PPP-Signalgebung umfassend analysieren zu können, ist es wichtig, die verschiedenen Holoenzyme in einer Zelle oder einem Gewebe zu untersuchen. Große Fortschritte wurden bei der Untersuchung des menschlichen Kinoms durch die Verwendung von Kinase-Inhibitor-Perlen erzielt, die als Multiplex-Inhibitor-Perlen oder Kinobeads bezeichnet werden, eine chemische Proteomstrategie, bei der Kinase-Inhibitoren auf Perlen immobilisiert werden und Massenspektrometrie verwendet wird, um angereicherte Kinasen und ihre Interaktorenzu identifizieren 10,11,12,13.
Wir haben einen ähnlichen Ansatz für das Studium der PPP-Biologie etabliert. Diese Technik beinhaltet die Affinitätserfassung von PPP-katalytischen Untereinheiten unter Verwendung von Perlen mit einem immobilisierten, nicht-selektiven PPP-Inhibitor namens Microcystin-LR (MCLR), der als Phosphatase-Inhibitor-Perlen (PIBs) bezeichnet wird14,15. Im Gegensatz zu anderen Methoden, die die endogene Markierung oder Expression exogener PPP-Untereinheiten erfordern, die die Proteinaktivität oder -lokalisation verändern könnten, ermöglicht PIB-MS die Anreicherung von endogenen PPP-katalytischen Untereinheiten, ihren assoziierten regulatorischen und gerüstbildenden Untereinheiten und interagierenden Proteinen (als PPPom bezeichnet) aus Zellen und Geweben zu einem bestimmten Zeitpunkt oder unter bestimmten Behandlungsbedingungen. MCLR hemmt PP1, PP2A, PP4-6, PPT und PPZ bei nanomolaren Konzentrationen, wodurch PIBs bei der Anreicherung für dasPPPome 16 hochwirksam sind. Diese Methode kann für den Einsatz auf jedem Ausgangsmaterial von Zellen bis hin zu klinischen Proben skaliert werden. Hier beschreiben wir detailliert den Einsatz von PIBs und Massenspektrometrie (PIB-MS), um das endogene PPPom und seine Modifikationszustände effizient zu erfassen, zu identifizieren und zu quantifizieren.
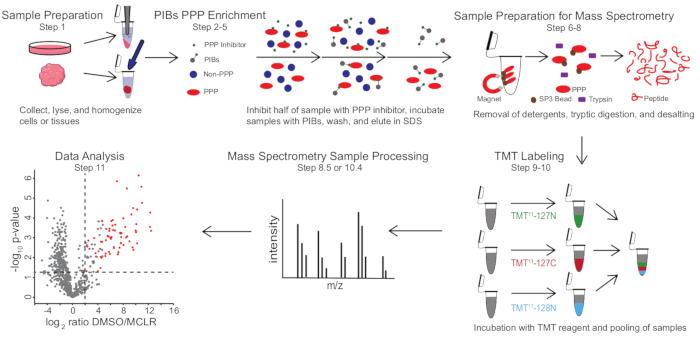
Abbildung 1: Visuelle Zusammenfassung des PIB-MS-Protokolls. In einem PIB-MS-Experiment können Proben in verschiedenen Formen gewonnen werden, von Zellen bis hin zu Tumoren. Die Probe wird vor der PPP-Anreicherung gesammelt, lysiert und homogenisiert. Zur Anreicherung für PPPs wird das Lysat mit PIBs mit oder ohne PPP-Inhibitor, wie MCLR, inkubiert. Die PIBs werden dann gewaschen und PPPs werden unter Denaturierungsbedingungen eluiert. Die Proben werden für die massenspektrometrische Analyse durch Entfernen von Reinigungsmitteln durch SP3-Proteinanreicherung, tryptische Verdauung und Entsalzung vorbereitet. Proben können dann vor der massenspektrometrischen Analyse optional TMT-markiert werden. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
PIB-MS umfasst die Lyse und Klärung von Zellen oder Geweben, die Inkubation des Lysats mit PIBs, die Elution und die Analyse des Eluats über westliche Blotting- oder Massenspektrometrie-basierte Ansätze (Abbildung 1). Die Zugabe von freiem MCLR kann als Steuerung verwendet werden, um spezifische PIB-Bindemittel von unspezifischen Interaktoren zu unterscheiden. Für die meisten Anwendungen kann ein markierungsfreier Ansatz verwendet werden, um Proteine in Eluaten direkt zu identifizieren. In Fällen, in denen eine genauere Quantifizierung oder die Identifizierung von Arten mit geringer Häufigkeit erforderlich ist, kann die Weiterverarbeitung mit Tandem-Massen-Tag-Kennzeichnung (TMT) verwendet werden, um die Abdeckung zu erhöhen und den Input zu verringern.
