يحتوي جسم الإنسان على ما يقدر بنحو 10-100 تريليون خلية ميكروبية حية (البكتيريا والفطريات القديمة) ، والتي توجد بشكل أساسي في الأمعاء والجلد والبيئات المخاطية1. في حالة صحية ، توفر هذه الفوائد لمضيفها ، بما في ذلك إنتاج الفيتامينات ، ونضج الجهاز المناعي ، وتحفيز الاستجابات المناعية الفطرية والتكيفية لمسببات الأمراض ، وتنظيم التمثيل الغذائي للدهون ، وتعديل استجابات الإجهاد ، وأكثر من ذلك ، مع التأثير على النمو والتطور ، وظهور المرض ، والشيخوخة2،3،4،5 . تتطور ميكروبات الأمعاء أيضا بشكل كبير طوال الحياة. يحدث التطور الأكثر جذرية خلال مرحلة الرضاعة والطفولة المبكرة6 ، ولكن تحدث تغييرات كبيرة أيضا مع تقدم العمر ، بما في ذلك انخفاض في وفرة Bifidobacterium وزيادة في Clostridium و Lactobacillus و Enterobacteriaceae و Enterococcus الأنواع 7. يمكن أن يؤدي نمط الحياة إلى تغيير تكوين ميكروبات الأمعاء مما يؤدي إلى dysbiosis (فقدان البكتيريا المفيدة ، فرط نمو البكتيريا الانتهازية) ، مما يؤدي إلى أمراض مختلفة مثل مرض التهاب الأمعاء والسكري والسمنة5 ، ولكن أيضا المساهمة في مرض الزهايمر وأمراض باركنسون8،9،10،11.
ساهم هذا الإدراك بشكل حاسم في تحسين مفهوم محور الأمعاء والدماغ (GBA) ، حيث تعتبر التفاعلات بين فسيولوجيا الأمعاء (بما في ذلك الآن الميكروبات الموجودة داخلها) والجهاز العصبي المنظم الرئيسي لعملية التمثيل الغذائي للحيوان والوظائف الفسيولوجية12. ومع ذلك ، فإن الدور الدقيق للميكروبات في إشارات الأمعاء والدماغ وآليات العمل المرتبطة بها بعيدة كل البعد عن الفهم الكامل13. مع كون ميكروبيوتا الأمعاء محددا رئيسيا للشيخوخة الصحية ، أصبحت كيفية تعديل البكتيريا لعملية الشيخوخة موضوعا للبحث المكثف والجدل6،14،15.
مع إثبات أن الدودة المستديرة Caenorhabditis elegans تستضيف ميكروبيوتا الأمعاء الصادقة التي تهيمن عليها – كما هو الحال في الأنواع الأخرى – من قبل Bacteroidetes و Firmicutes و Actinobacteria16،17،18،19،20 ، صعودها السريع كمنصة تجريبية لدراسة التفاعلات المشتركة بين المضيفوالأمعاء 21،22،23،24 ، 25،26 وسعت بشكل كبير ترسانة التحقيقلدينا 26،27،28،29. على وجه الخصوص ، يمكن تكييف الأساليب التجريبية عالية الإنتاجية المتاحة ل C. elegans لدراسة النظام الغذائي الجيني ، والتفاعلات الجينية الدوائية ، والممرض الجيني ، وما إلى ذلك ، لاستكشاف كيفية تأثير العزلات البكتيرية والكوكتيلات على صحة C. elegans وشيخوختها بسرعة.
يصف البروتوكول الحالي خط أنابيب تجريبي لفحص صفائف من العزلات البكتيرية أو المخاليط الموضوعة في ألواح متعددة الآبار للتأثيرات على مقاومة الإجهاد C. elegans كبديل للصحة ، والتي يمكن استخدامها لتحديد البروبيوتيك. يوضح بالتفصيل كيفية تنمية أعداد كبيرة من الديدان والتعامل مع المصفوفات البكتيرية في تنسيقات ألواح 96 و 384 بئرا قبل معالجة الديدان لتحليل مقاومة الإجهاد الآلي باستخدام قارئ لوحة مضان (الشكل 1). يعتمد هذا النهج على فحوصات البقاء الآلية الخالية من الملصقات (LFASS)30 التي تستغل ظاهرة مضان الموت31 ، حيث تنتج الديدان المحتضرة دفعة من التألق الأزرق الذي يمكن استخدامه لتحديد وقت الوفاة. ينبعث التألق الأزرق بواسطة استرات الجلوكوزيل لحمض الأنثرانيليك المخزن في حبيبات الأمعاء C. elegans (نوع من العضيات المرتبطة بالليزوزوم) ، والتي تنفجر عندما يتم تشغيل شلال في أمعاء الدودة عند الوفاة31.
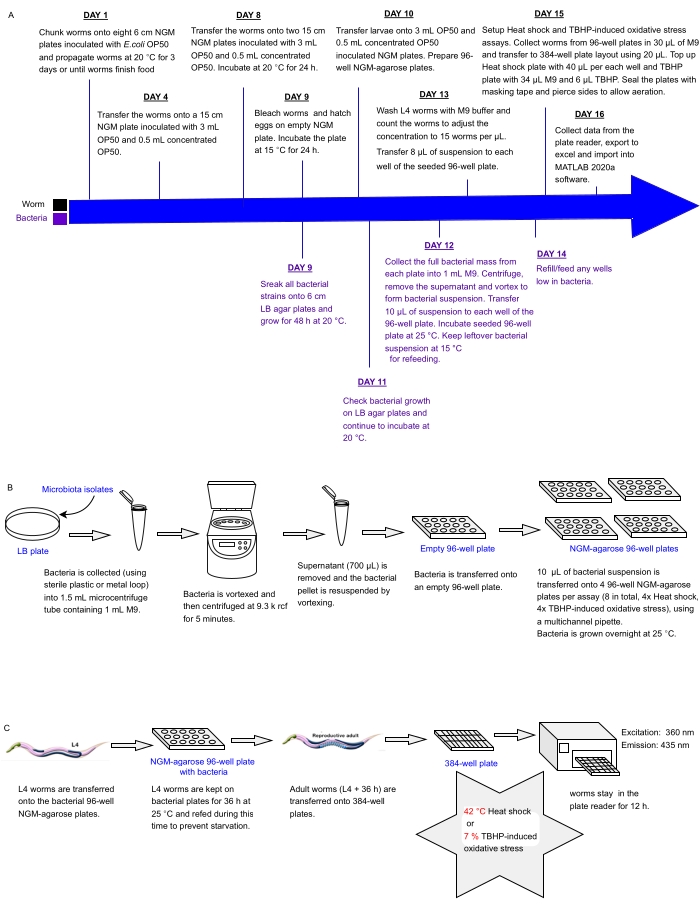
الشكل 1: سير العمل التجريبي للفحص عالي الإنتاجية للعزلات البكتيرية مع التأثير على مقاومة C. elegans للإجهاد . (أ) الجدول الزمني لصيانة الديدان والبكتيريا وإعداد المقايسة. (ب) إعداد صفيف صفيحة بكتيرية 96 بئرا والتعامل معها. (ج) إعداد لوحة دودة 384 بئرا. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
