Il corpo umano ospita circa 10-100 trilioni di cellule microbiche vive (batteri, funghi archaea), che si trovano principalmente negli ambienti dell’intestino, della pelle e delle mucose1. In uno stato sano, questi forniscono benefici al loro ospite, tra cui la produzione di vitamine, la maturazione del sistema immunitario, la stimolazione delle risposte immunitarie innate e adattative agli agenti patogeni, la regolazione del metabolismo dei grassi, la modulazione delle risposte allo stress e altro ancora, con un impatto sulla crescita e lo sviluppo, l’insorgenza della malattia e l’invecchiamento 2,3,4,5 . Anche il microbiota intestinale si evolve considerevolmente nel corso della vita. L’evoluzione più drastica si verifica durante l’infanzia e la prima infanzia6, ma cambiamenti significativi si verificano anche con l’età, tra cui una diminuzione dell’abbondanza di Bifidobacterium e un aumento di Clostridium, Lactobacillus, Enterobacteriaceae e Enterococcus specie7. Lo stile di vita può alterare ulteriormente la composizione microbica intestinale portando alla disbiosi (perdita di batteri benefici, crescita eccessiva di batteri opportunisti), causando varie patologie come malattie infiammatorie intestinali, diabete e obesità5, ma anche contribuendo alle malattie di Alzheimer e Parkinson 8,9,10,11.
Questa consapevolezza ha contribuito in modo critico ad affinare il concetto di asse intestino-cervello (GBA), dove le interazioni tra la fisiologia intestinale (ora inclusi i microbi al suo interno) e il sistema nervoso sono considerate il principale regolatore del metabolismo animale e delle funzioni fisiologiche12. Tuttavia, il ruolo preciso del microbiota nella segnalazione intestino-cervello e i meccanismi d’azione associati sono lontani dall’essere pienamente compresi13. Con il microbiota intestinale che è un determinante chiave dell’invecchiamento sano, il modo in cui i batteri modulano il processo di invecchiamento è diventato oggetto di intense ricerche e polemiche 6,14,15.
Con la dimostrazione che il nematode Caenorhabditis elegans ospita un microbiota intestinale in buona fede dominato – come in altre specie – da Bacteroidetes, Firmicutes e Actinobacteria 16,17,18,19,20, la sua rapida ascesa come piattaforma sperimentale per studiare le interazioni commensali ospite-intestino21,22,23,24 ,25,26 ha notevolmente ampliato il nostro arsenale investigativo26,27,28,29. In particolare, gli approcci sperimentali ad alto rendimento disponibili per C. elegans per studiare le interazioni gene-dieta, gene-farmaco, gene-patogeno, ecc., possono essere adattati per esplorare rapidamente come isolati e cocktail batterici influenzano la salute e l’invecchiamento di C. elegans.
Il presente protocollo descrive una pipeline sperimentale per lo screening simultaneo di array di isolati o miscele batteriche impostati in piastre multipozzetto per gli effetti sulla resistenza allo stress di C. elegans come proxy per la salute, che può essere utilizzato per identificare i probiotici. Descrive in dettaglio come far crescere grandi popolazioni di vermi e gestire array batterici in formati di piastre a 96 e 384 pozzetti prima di elaborare i vermi per l’analisi automatizzata della resistenza allo stress utilizzando un lettore di piastre a fluorescenza (Figura 1). L’approccio si basa su saggi di sopravvivenza automatizzati label-free (LFASS)30 che sfruttano il fenomeno della fluorescenza di morte31, per cui i vermi morenti producono un’esplosione di fluorescenza blu che può essere utilizzata per individuare il momento della morte. La fluorescenza blu è emessa dagli esteri glucosilici dell’acido antranilico immagazzinati nei granuli intestinali di C. elegans (un tipo di organello correlato al lisosoma), che scoppiano quando una cascata necrotica viene innescata nell’intestino del verme alla morte31.
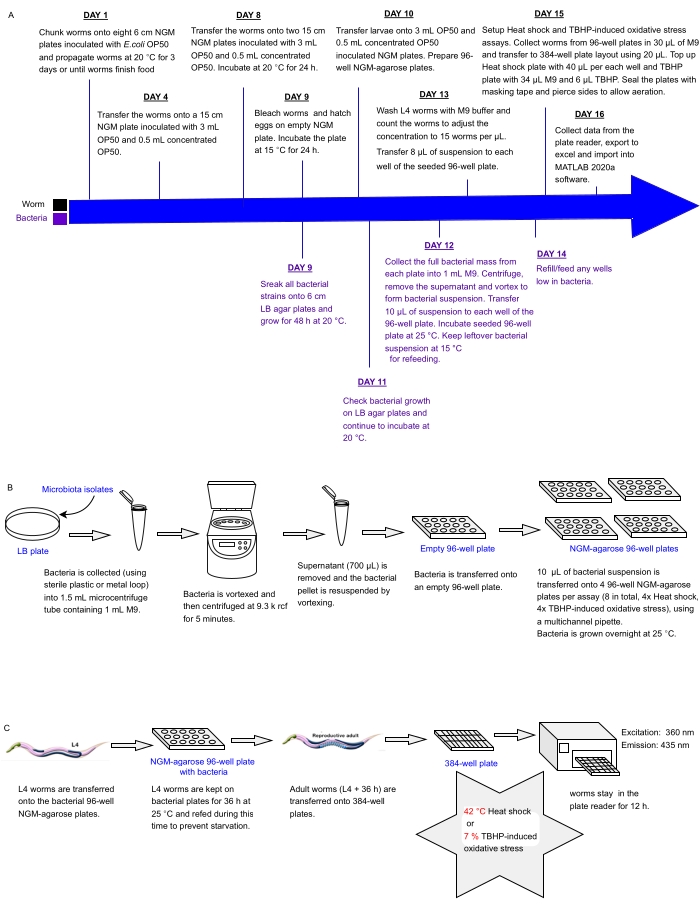
Figura 1: Flusso di lavoro sperimentale per lo screening ad alta produttività di isolati batterici con impatto sulla resistenza allo stress di C. elegans . (A) Cronologia per la manutenzione di vermi e batteri e la configurazione del test. (B) Configurazione e gestione dell’array di piastre batteriche a 96 pozzetti. (C) Configurazione della piastra a vite senza fine a 384 pozzetti. Fare clic qui per visualizzare una versione ingrandita di questa figura.
