Le corps humain abrite environ 10 à 100 billions de cellules microbiennes vivantes (bactéries, champignons archées), qui se trouvent principalement dans les environnements intestinaux, cutanés et muqueux1. Dans un état sain, ceux-ci offrent des avantages à leur hôte, y compris la production de vitamines, la maturation du système immunitaire, la stimulation des réponses immunitaires innées et adaptatives aux agents pathogènes, la régulation du métabolisme des graisses, la modulation des réponses au stress, etc., avec un impact sur la croissance et le développement, l’apparition de la maladie et le vieillissement 2,3,4,5 . Le microbiote intestinal évolue également considérablement tout au long de la vie. L’évolution la plus radicale se produit pendant la petite enfance et la petite enfance6, mais des changements significatifs se produisent également avec l’âge, notamment une diminution de l’abondance de Bifidobacterium et une augmentation des espèces de Clostridium, Lactobacillus, Enterobacteriaceae et Enterococcus 7. Le mode de vie peut altérer davantage la composition microbienne intestinale conduisant à une dysbiose (perte de bactéries bénéfiques, prolifération de bactéries opportunistes), entraînant diverses pathologies telles que les maladies inflammatoires de l’intestin, le diabète et l’obésité5, mais contribuant également aux maladies d’Alzheimer et de Parkinson 8,9,10,11.
Cette prise de conscience a contribué de manière critique à affiner le concept de l’axe intestin-cerveau (ACS), où les interactions entre la physiologie intestinale (y compris maintenant les microbes qu’elle contient) et le système nerveux sont considérées comme le principal régulateur du métabolisme animal et des fonctions physiologiques12. Cependant, le rôle précis du microbiote dans la signalisation intestin-cerveau et les mécanismes d’action associés sont loin d’être entièrement compris13. Le microbiote intestinal étant un déterminant clé du vieillissement en bonne santé, la façon dont les bactéries modulent le processus de vieillissement est devenue un sujet de recherche intense et de controverse 6,14,15.
Avec la démonstration que le ver rond Caenorhabditis elegans héberge un véritable microbiote intestinal dominé – comme chez d’autres espèces – par Bacteroidetes, Firmicutes, et Actinobacteria 16,17,18,19,20, son ascension rapide en tant que plate-forme expérimentale pour étudier les interactions commensales hôte-intestin21,22,23,24 ,25,26 a considérablement élargi notre arsenal d’enquête26,27,28,29. En particulier, les approches expérimentales à haut débit disponibles pour C. elegans pour étudier les interactions gène-alimentation, gène-médicament, gène-pathogène, etc., peuvent être adaptées pour explorer rapidement l’impact des isolats et des cocktails bactériens sur la santé et le vieillissement de C. elegans.
Le présent protocole décrit un pipeline expérimental permettant de dépister immédiatement des réseaux d’isolats ou de mélanges bactériens placés dans des plaques multipuits pour déterminer les effets sur la résistance au stress de C. elegans en tant qu’indicateur de la santé, ce qui peut être utilisé pour identifier les probiotiques. Il explique comment développer de grandes populations de vers et manipuler des réseaux bactériens dans des formats de plaques à 96 et 384 puits avant de traiter les vers pour une analyse automatisée de la résistance au stress à l’aide d’un lecteur de plaque de fluorescence (Figure 1). L’approche est basée sur des tests de survie automatisés sans marquage (LFASS)30 qui exploitent le phénomène de fluorescence de mort31, par lequel les vers mourants produisent une explosion de fluorescence bleue qui peut être utilisée pour déterminer le moment de la mort. La fluorescence bleue est émise par les esters glucosyles de l’acide anthranilique stockés dans les granules intestinaux de C. elegans (un type d’organite apparenté au lysosome), qui éclatent lorsqu’une cascade nécrotique est déclenchée dans l’intestin du ver à la mort31.
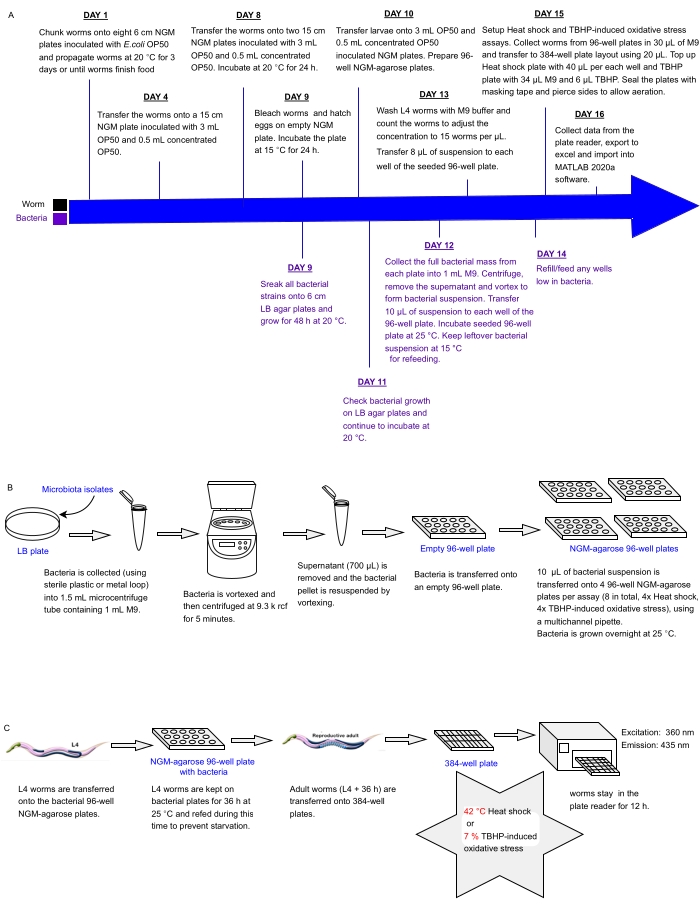
Figure 1 : Flux de travail expérimental pour le criblage à haut débit d’isolats bactériens ayant un impact sur la résistance de C. elegans au stress. (A) Calendrier pour l’entretien et la mise en place des tests vers et bactériens. (B) Installation et manipulation de réseaux de plaques bactériennes à 96 puits. (C) Installation de plaques à vis sans fin à 384 puits. Veuillez cliquer ici pour voir une version agrandie de cette figure.
