Människokroppen har uppskattningsvis 10-100 biljoner levande mikrobiella celler (bakterier, archaea-svampar), som främst finns i tarm-, hud- och slemhinnemiljöerna1. I ett hälsosamt tillstånd ger dessa fördelar för sin värd, inklusive vitaminproduktion, mognad av immunsystemet, stimulering av medfödda och adaptiva immunsvar mot patogener, reglering av fettmetabolism, modulering av stressreaktioner och mer, med inverkan på tillväxt och utveckling, sjukdomsdebut och åldrande 2,3,4,5 . Tarmmikrobiotan utvecklas också avsevärt under hela livet. Den mest drastiska utvecklingen sker under spädbarn och tidig barndom6, men betydande förändringar sker också med åldern, inklusive en minskning av Bifidobacterium överflöd och en ökning av Clostridium, Lactobacillus, Enterobacteriaceae, och Enterococcus arter7. Livsstil kan ytterligare förändra tarmens mikrobiella sammansättning som leder till dysbios (förlust av fördelaktiga bakterier, överväxt av opportunistiska bakterier), vilket resulterar i olika patologier som inflammatorisk tarmsjukdom, diabetes och fetma5, men också bidrar till Alzheimers och Parkinsons sjukdomar 8,9,10,11.
Denna insikt har kritiskt bidragit till att förfina begreppet tarm-hjärnaxeln (GBA), där interaktioner mellan tarmfysiologi (nu inklusive mikroberna i den) och nervsystemet anses vara den viktigaste regulatorn för djurmetabolism och fysiologiska funktioner12. Emellertid, den exakta rollen av mikrobiota i tarm-hjärna signalering och de associerade verkningsmekanismerna är långt ifrån helt förstådd13. Med tarmmikrobiota som en viktig determinant för hälsosamt åldrande har hur bakterier modulerar åldringsprocessen blivit föremål för intensiv forskning och kontrovers 6,14,15.
Med demonstrationen att rundmask Caenorhabditis elegans är värd för en bonafide tarmmikrobiota dominerad-som i andra arter-av Bacteroidetes, Firmicutes och Actinobacteria 16,17,18,19,20, dess snabba uppgång som en experimentell plattform för att studera värd-tarm kommensala interaktioner 21,22,23,24 ,25,26 har avsevärt utökat vår utredningsarsenal26,27,28,29. I synnerhet kan experimentella metoder med hög genomströmning tillgängliga för C. elegans för att studera gen-diet, gen-läkemedel, gen-patogen, etc. interaktioner, anpassas för att snabbt utforska hur bakterieisolat och cocktails påverkar C. elegans hälsa och åldrande.
Detta protokoll beskriver en experimentell pipeline för att på en gång screena arrays av bakterieisolat eller blandningar som sätts i multiwell plattor för effekter på C. elegans stressresistens som en proxy för hälsa, som kan användas för att identifiera probiotika. Den beskriver hur man odlar stora maskpopulationer och hanterar bakteriella arrayer i 96- och 384-brunnsplattformat innan man bearbetar maskar för automatiserad stressmotståndsanalys med hjälp av en fluorescensplattläsare (figur 1). Tillvägagångssättet bygger på etikettfria automatiserade överlevnadsanalyser (LFASS)30 som utnyttjar fenomenet dödsfluorescens31, där döende maskar producerar en explosion av blå fluorescens som kan användas för att fastställa dödstiden. Blå fluorescens avges av glukosylestrar av anthranilsyra lagrade i C. elegans tarmgranulat (en typ av lysosomrelaterad organell), som spricker när en nekrotisk kaskad utlöses i masktarmen vid döden31.
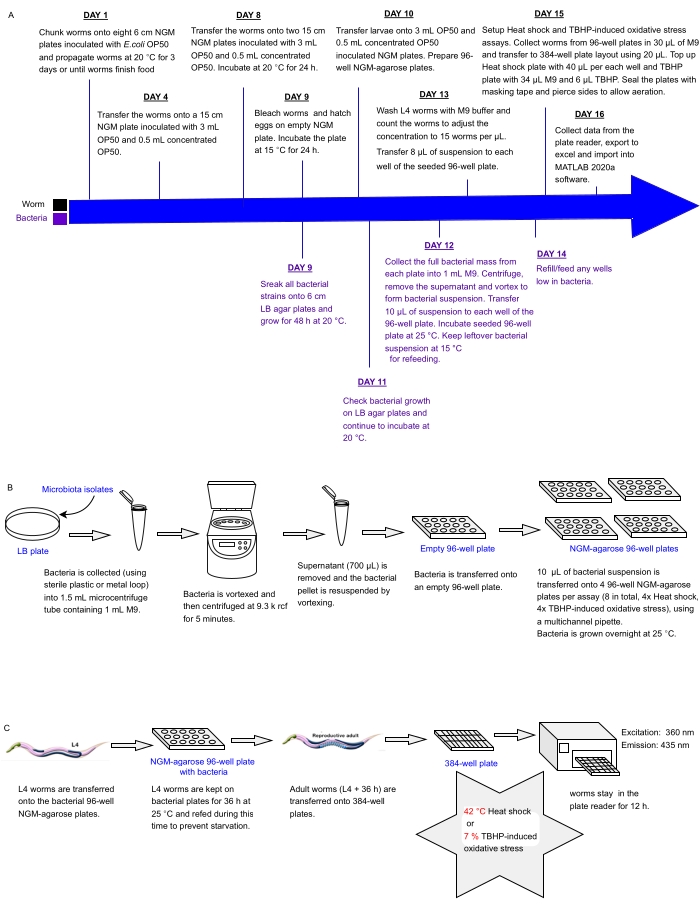
Figur 1: Experimentellt arbetsflöde för screening med hög genomströmning av bakterieisolat med inverkan på C. elegans motståndskraft mot stress . (A) Tidslinje för underhåll och analys av mask och bakterier. (B) Installation och hantering av bakterieplattor med 96 brunnar. (C) Installation av maskplattor med 384 brunnar. Klicka här för att se en större version av denna siffra.
