骨格筋における筋肉内脂肪形成とその細胞起源を調べるためのガイド
Summary
健康な筋肉組織を筋肉内脂肪で置き換えることは、ヒトの疾患および状態の顕著な特徴である。このプロトコルは、筋肉内脂肪を視覚化、画像化、定量化する方法を概説し、筋肉内脂肪形成の根底にあるメカニズムの厳密な研究を可能にします。
Abstract
線維脂肪原性前駆細胞(FAP)は、骨格筋の恒常性および再生中に重要な役割を果たす間葉間質細胞である。FAPは、分子筋繊維足場として作用する細胞外マトリックスを構築し、維持する。さらに、FAPは筋幹細胞(MuSC)によって感知される多数の有益な因子を分泌するため、筋線維再生に不可欠です。しかし、罹患状態では、FAPは筋肉内脂肪および線維性瘢痕組織の細胞起源である。この脂肪線維症は、サルコペニアおよびデュシェンヌ型筋ジストロフィーなどの神経筋疾患の特徴である。FAPが筋肉内脂肪に分化する理由と方法を決定する際の重要な障壁の1つは、特に凍結組織切片における脂肪細胞の効果的な保存とその後の視覚化です。スナップ凍結などの骨格筋組織処理の従来の方法は、個々の脂肪細胞の形態を適切に保存せず、それによって正確な視覚化および定量化を妨げる。このハードルを克服するために、骨格筋切片の脂肪細胞形態を保存し、筋肉内脂肪の視覚化、イメージング、定量化を可能にする厳格なプロトコルが開発されました。このプロトコルはまた、RT-qPCRのために筋肉組織の一部を処理する方法を概説し、脂肪生成遺伝子の発現の違いを見ることによって、観察された脂肪形成の変化を確認することを可能にする。さらに、筋肉サンプルの全マウント免疫蛍光によって脂肪細胞を視覚化するように適合させることができる。最後に、このプロトコルは、FAPの脂肪生成変換を研究するために 、Pdgfrα発現FAPの遺伝的系統追跡を実行する方法を概説する。このプロトコルは、RT-qPCRによる確認とともに、脂肪細胞の高解像度で形態学的に正確な免疫蛍光画像を一貫して生成し、筋肉内脂肪の堅牢で厳密で再現性のある視覚化と定量を可能にします。ここで説明する分析パイプラインは、FAPが筋肉内脂肪にどのように分化するかについての理解を深めるための最初のステップであり、脂肪形成を防ぐための新しい介入を検証するためのフレームワークを提供します。
Introduction
脂肪線維症を伴う健康な筋肉組織の浸潤は、デュシェンヌ型筋ジストロフィー(DMD)および他の神経筋疾患、ならびにサルコペニア、肥満、および糖尿病の顕著な特徴である1,2,3,4,5,6,7,8,9,10 .これらの状態における脂肪浸潤の増加は筋肉機能の低下と強く関連していますが、筋肉内脂肪が形成される理由と方法に関する私たちの知識はまだ限られています。FAPsは、骨格筋を含むほとんどの成人器官に存在する多能性間葉間質細胞集団である11、12。しかし、加齢および慢性疾患において、FAPは線維性瘢痕組織を産生し、脂肪細胞に分化し、個々の筋線維の間に位置し、筋肉内脂肪を形成する13、14、15、16、17、18、19、20を形成する。
筋肉内脂肪形成との戦いを開始するには、FAPが脂肪細胞にどのように変化するかのメカニズムを定義する必要があります。PDGFRαは、複数の種13、16、17、18、20、21、22、23、24、25、26、27の筋肉内のFAPを同定するための分野における「ゴールドスタンダード」マーカーである。その結果、Pdgfrαプロモーターの制御下で、いくつかのマウスタモキシフェン誘導性Cre系統が生成され、Cre-LoxP系を用いてインビボでFAPを遺伝的に操作することが可能になった27、28、29。例えば、この誘導性Cre系統を遺伝子レポーターと組み合わせることにより、FAPsの系統追跡を行うことができ、この戦略を筋肉および白色脂肪組織におけるFAPsの運命マップに適用することに成功した20,30。リネージュトレースに加えて、これらのCreラインはFAPから脂肪への変換を研究するための貴重なツールを提供します。
FAPの筋肉内脂肪への脂肪生成変換のメカニズムを定義する際の1つの大きな障害は、異なる条件下で形成された筋肉内脂肪の量を厳密かつ再現性よく定量する能力である。重要なのは、筋肉と脂肪組織の保存のバランスを取り、これを脂肪細胞を視覚化するための利用可能な染色方法と一致させることです。例えば、骨格筋はしばしば事前の固定なしにスナップ凍結され、筋線維は保存されるが脂肪細胞の形態を破壊する(図1)。対照的に、固定に続いてパラフィン包埋を行うと、脂肪細胞を含む最良の組織組織学を表示しながら、すべての脂質を除去し、それによって一般的に使用される色素オイルレッドOなどのほとんどの親油性色素を使用不能にする。
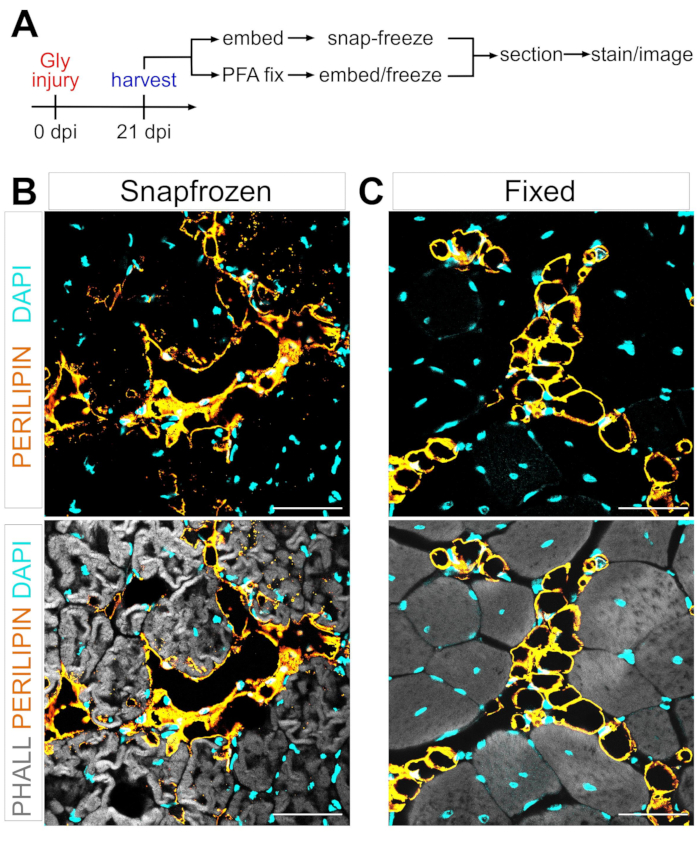
図1:スナップ凍結した筋肉組織と固定された筋肉組織における筋肉内脂肪の代表的な画像。グリセロール損傷後21日目に脂肪細胞(黄色)、筋線維(灰色)、および核(シアン)内の(B)スナップフリーズおよび(C)固定TAsの両方を示す免疫蛍光画像。スケール バー: 50 μm。この図の拡大版を表示するには、ここをクリックしてください。
ここで説明するプロトコルは、筋線維および脂肪細胞の形態を保持し、複数の細胞型の視覚化および分析を可能にする。このアプローチは、パラホルムアルデヒド(PFA)固定筋肉組織における脂肪細胞の免疫蛍光染色に基づいており、複数の抗体との共染色を可能にします。また、ホールマウントイメージングを使用して、無傷の組織中の筋肉内脂肪を空間的に表示するように容易に適合させることができ、それによって筋肉内の脂肪の細胞微小環境に関する情報を提供する。加えて、このプロトコルは、筋肉の健康状態を評価するための重要な測定値である固定筋肉組織31における筋線維の断面積を決定するために、最近公表された我々のアプローチと組み合わせることができる。このアプローチを遺伝的系統追跡と組み合わせて、脂肪細胞へのFAPの分化を運命マップすることもここで概説されています。したがって、ここで説明する汎用性の高いプロトコルは、組織切片および無傷の組織におけるFAPおよび筋肉内脂肪へのそれらの分化の厳密かつ再現性のある評価を可能にする。
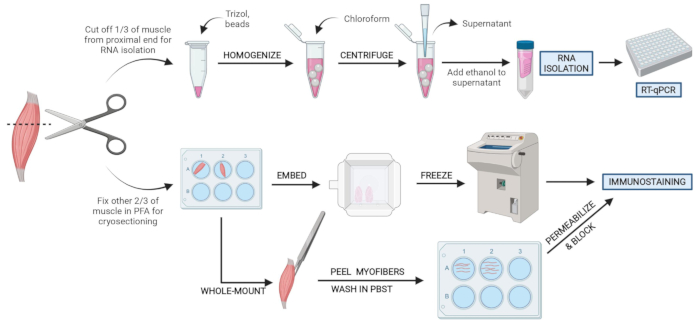
図2:概略プロトコルの概要 TAの3分の1を除去し、スナップフリーズし、ホモジナイズして、その後のRNA単離およびRT-qPCR を介した 転写解析を行う組織処理の概略概要。TAの他の3分の2はPFA固定され、凍結切片または全マウント繊維の免疫染色のために処理されます。 この図の拡大版を表示するには、ここをクリックしてください。
Protocol
Representative Results
Discussion
このプロトコルは、筋肉内脂肪の効率的な視覚化と厳密な定量化を可能にする広範で詳細なプロトコルの概要を示しています。同じ筋肉を2つの部分に分割し、1つは免疫蛍光に、もう1つはRT-qPCR分析に使用することで、このプロトコルも非常に汎用性があります。また、FAPの遺伝的系統追跡と組み合わせて、特定の条件下で脂肪細胞への変換を研究することもでき、複数の追加の細胞型を標識および画像化するために高度に適応可能である。
筋肉内脂肪を視覚化するために最も一般的に使用される方法は、パラフィン切片に続いてヘマトキシリンおよびエオジン染色、またはオイルレッドO(ORO)などの親油性染料で染色された凍結切片である。しかし、パラフィン処理された組織は最高の組織学を維持する一方で、同じプロセスはまた、親油性染料の使用を妨げるすべての脂質を抽出する。親油性染色法はPFA固定組織切片と非固定組織切片の両方に作用するが、脂肪滴はカバースリップに圧力を加えることによって容易に変位し、それによって筋肉内脂肪の空間分布を歪める。これを回避するために、最近の研究では、ホールマウントアプローチを使用してORO+脂肪細胞を視覚化するための厳格なプロトコルが確立されました。このために、著者らはTAを脱細胞化し、TA41全体にわたる筋肉内脂肪の空間分布を視覚化した。この技術と同じくらい強力ですが、それはまた、追加の細胞構造をマークするために他の共染色剤の使用を防ぎます。ここで紹介するホールマウント免疫蛍光アプローチは、脂肪細胞をさまざまなマーカーと共染色するために使用でき、細胞環境の微細なマッピングを可能にします。しかし、1つの大きな課題は、抗体の組織浸透である。より多くの繊維が一緒に保たれるほど、抗体が均等に浸透し、利用可能なすべての抗原に結合することがより困難になります。したがって、この方法は、繊維の小さなグループを見るときに最も効果的です。同時に、筋肉内脂肪の全体的な解剖学的位置は、小さな剥離した繊維束のみに焦点を当てると失われているため、これも制限です。しかし、新しい組織クリアリング方法と新しいイメージング技術の現在の開発により、将来的には組織の浸透と視覚化が可能になります42,43,44。
筋肉組織の事前固定は脂肪細胞の形態を維持する一方で、筋肉の健康の重要な測定値である筋線維のサイズを評価するという課題も生み出します。ミオファイバーサイズは、ミオファイバーの断面積を測定することにより求められる。我々は以前、筋肉組織の事前固定により、筋線維の輪郭を描くために利用可能なほとんどのマーカーが失敗することを報告した31。このハードルを克服するために、我々は、固定筋肉切片31においても筋線維サイズの測定を可能にする新規な画像セグメンテーションパイプラインを開発した。したがって、このプロトコルと組み合わせて、筋肉組織の事前固定によって引き起こされるほとんどの欠点を克服する堅牢で効率的な組織処理パイプラインを確立しました。
このアプローチのもう1つの大きな利点は、汎用性です。TAを2つの部分に分割することで、1つの筋肉から得られる情報量が最大化されます。これは動物の数を減らすだけでなく、遺伝子発現を通して組織学を確認することによって、そしてその逆もまた然りであることによって、制御の余分な層を追加します。さらに、脂肪生成遺伝子を超えて多くの異なる遺伝子を調べることができる。単離されたRNAは、筋肉全体のRNAseq実験にも使用することができる。最後に、スナップ凍結筋肉片は、タンパク質作業にも使用できます。このプロトコルの1つの制限は、傷害がTAの全長にわたって一貫していない可能性である。これは、2つの筋肉部分がそれらに含まれる筋肉内脂肪の量で発散するシナリオにつながる可能性があり、下流の分析からそのようなサンプルを除外することを正当化する可能性がある。したがって、筋肉内脂肪の量に関する主要な結論を引き出すために単にRT-qPCRに頼るのではなく、むしろ組織学的定量化を支持するデータとして行うことが推奨される。
一緒に、このプロトコルは、脂肪線維症と戦うための新しい治療選択肢を開発するための最初のステップである筋肉内脂肪の視覚化と定量化を可能にする、堅牢で効率的で厳格な組織処理パイプラインを概説します。同時に、それは汎用性があり、筋肉内の多くの異なる細胞型ならびに他の組織の脂肪細胞に適応させることができる。
Disclosures
The authors have nothing to disclose.
Acknowledgements
コピンケ研究所のメンバーには、データ収集と原稿の批判的読解を手伝ってくれたことに感謝します。また、フロリダ大学の筋学研究所のメンバーにも、原稿に関する貴重な意見に感謝します。この作業は、NIH助成金1R01AR079449によって支援されました。図 2 は Biorender で作成したものです。
Materials
| 16% PFA (Pack of 12, 10 mL bottles) | Electron Miscroscopy Sciences | 15710 | |
| 2.0 mL Microcentrifuge Tubes | Fisher Scientific | 05-408-138 | microcentrifuge tubes for snapfreezing/bead beating |
| 2-Methylbutane (4 L) | Fisher Chemical | O3551-4 | isopentane |
| Absolute Ethanol (200 proof) | ThermoFisher Scientific | BP2818100 | |
| AffiniPure Fab fragment donkey anti-mouse | Jackson ImmunoResearch | 715-007-003 | mouse-on-mouse blocking |
| Alexa Fluor 488 donkey anti-chicken secondary antibody | Jackson ImmunoResearch | 703-545-155 | |
| Alexa Fluor 488 donkey anti-mouse secondary antibody | Invitrogen | A21202 | |
| Alexa Fluor 488 donkey anti-rabbit secondary antibody | Invitrogen | A21206 | |
| Alexa Fluor 568 donkey anti-goat secondary antibody | Invitrogen | A11057 | |
| Alexa Fluor 568 donkey anti-rabbit secondary antibody | Invitrogen | A11037 | |
| Alexa Fluor 568 Phalloidin antibody | Invitrogen | A12380 | Dissolved in 1.5 mL methanol (~66 µM working solution) |
| BioLite 24-well Multidishes | ThermoFisher Scientific | 930186 | 24 well plate for PFA tissue incubation |
| Biometra TOne | analytikjena | 8462070301 | Thermal cycler |
| Chicken anti-GFP antibody | Aves Labs | GFP-1020 | |
| Chloroform/isoamyl alcohol 24:1(v/v) for molecular biology, DNAse, RNAse, and Protease free | ThermoFisher Scientific | AC327155000 | |
| Corn oil | Sigma Aldrich | C8267 | |
| DAPI stain | Invitrogen | D1306 | 150 µM working solution in dH2O |
| Donkey Serum (100 mL) | Millipore Sigma | 5058837 | for blocking solution |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | sharp-tipped tweezers |
| Fine Scissors Straight 9 cm | Fine Science Tools | 14060-09 | |
| Fluoromount-G | SouthernBiotech | 0100-01 | mounting medium |
| Glycerol, 99.5%, for molecular biology (500 mL) | Acros Organics | 327255000 | |
| Goat anti-PDGFRα antibody | R&D | AF1062 | |
| Hybridization Oven | VWR | 230301V | for Tamoxifen incubation |
| ImmEdge Hydrophobic Barrier PAP Pen | Vector Laboratories | H-4000 | hydrophobic pen |
| Insulin Syringe with Micro-Fine IV needle (28 G) | BD | 329461 | |
| Insulin Syringe with Slip Tip, 1 mL | BD | 329654 | Insulin syringe without needle, for oral gavaging |
| iScript cDNA Synthesis Kit | Bio-Rad | 1708890 | |
| Isoflurane | Patterson Veterinary | 78938441 | |
| Leica DMi8 inverted microscope | Leica | micrscope used for widefield IF and confocal imaging | |
| Micro Slides | VWR | 48311-703 | positively charged microscope slides |
| mouse anti-MYOD antibody | Invitrogen | MA1-41017 | |
| mouse anti-PAX7 antibody (supernatant) | DSHB | AB 428528 | |
| MX35 Premier+ Microtome blades | ThermoFisher Scientific | 3052835 | microtome blades |
| NanoDrop 2000 Spectrophotometer | ThermoFisher Scientific | ND2000 | spectrophotometer for RNA yield |
| Play-Doh | Hasbro | modeling compound | |
| PowerUp SYBR Green Master Mix | ThermoFisher Scientific | A25742 | green dye PCR master mix |
| Puralube Vet Ointment | Puralube | 17033-211-38 | vet ophthalmic ointment |
| QuantStudi 6 Flex Real-Time 384-well PCR System | Applied Biosystems | 4485694 | qPCR machine |
| Rabbit anti-perilipin antibody | Cell Signaling Technology | 9349S | |
| Red-Rotor Shaker | Hoefer Scientific | PR70-115V | shaker for IF staining |
| Richard-Allan Scientific Slip-Rite Cover Glass | ThermoFisher Scientific | 152460 | coverslips |
| RNeasy Mini Kit | QIAGEN | 74106 | contains mini spin columns |
| Safe-Lock Tubes 1.5 ml, natural | Eppendorf | 22363204 | |
| Sample Tubes RB (2 mL) | QIAGEN | 990381 | |
| Sodium azide | Alfa Aesar | 14314 | |
| Stainless Steel Beads, 2.8 mm | Precellys | KT03961-1-101.BK | small beads |
| Stainless Steel Beads, 5 mm | QIAGEN | 69989 | medium beads |
| Stainless Steel Beads, 7 mm | QIAGEN | 69990 | large beads |
| Stainless Steel Disposable Scalpels | Miltex | 327-4102 | scalpel |
| Stainless steel feeding tube, 20 G x 38 mm, straight | Instech Laboratories | FTSS-20S-3 | gavage needle |
| Tamoxifen | Toronto Research Chemicals | T006000 | |
| Tissue Plus O.C.T. Compound | Fisher HealthCare | 4585 | embedding medium |
| TissueLyser LT | QIAGEN | 85600 | bead beater |
| TissueLyser LT Adapter, 12-Tube | QIAGEN | 69980 | |
| Tissue-Tek Cryomold | Sakura | 4566 | specimen molds |
| Triton X-100 | Alfa Aesar | A16046 | |
| TRIzol Reagent | ThermoFisher Scientific | 15596026 | guanidium thiocyanate |
| Tween20 (500 mL) | Fisher BioReagents | BP337-500 | |
| VWR Micro Slides – Superfrost Plus | VWR | 48311703 | |
| Wheaton Coplin staining jars | Millipore Sigma | S6016 | Coplin jar |
References
- Milad, N., et al. Increased plasma lipid levels exacerbate muscle pathology in the mdx mouse model of Duchenne muscular dystrophy. Skeletal Muscle. 7 (1), 19 (2017).
- Goodpaster, B. H., et al. Obesity, regional body fat distribution, and the metabolic syndrome in older men and women. Archives of Internal Medicine. 165 (7), 777-783 (2005).
- Goodpaster, B. H., et al. Association between regional adipose tissue distribution and both type 2 diabetes and impaired glucose tolerance in elderly men and women. Diabetes Care. 26 (2), 372-379 (2003).
- Goodpaster, B. H., et al. The loss of skeletal muscle strength, mass, and quality in older adults: the health, aging and body composition study. The Journals of Gerontology, Series A: Biological Sciences and Medical Sciences. 61 (10), 1059-1064 (2006).
- Goodpaster, B. H., Thaete, F. L., Kelley, D. E. Thigh adipose tissue distribution is associated with insulin resistance in obesity and in type 2 diabetes mellitus. American Journal of Clinical Nutrition. 71 (4), 885-892 (2000).
- Goodpaster, B. H., Theriault, R., Watkins, S. C., Kelley, D. E. Intramuscular lipid content is increased in obesity and decreased by weight loss. Metabolism. 49 (4), 467-472 (2000).
- Burakiewicz, J., et al. Quantifying fat replacement of muscle by quantitative MRI in muscular dystrophy. Journal of Neurology. 264 (10), 2053-2067 (2017).
- Murphy, W. A., Totty, W. G., Carroll, J. E. MRI of normal and pathologic skeletal muscle. American Journal of Roentgenology. 146 (3), 565-574 (1986).
- Willcocks, R. J., et al. Multicenter prospective longitudinal study of magnetic resonance biomarkers in a large duchenne muscular dystrophy cohort. Annals of Neurology. 79 (4), 535-547 (2016).
- Wokke, B. H., et al. Quantitative MRI and strength measurements in the assessment of muscle quality in Duchenne muscular dystrophy. Neuromuscular Disorders. 24 (5), 409-416 (2014).
- Contreras, O., Rossi, F. M. V., Theret, M. Origins, potency, and heterogeneity of skeletal muscle fibro-adipogenic progenitors-time for new definitions. Skeletal Muscle. 11 (1), 16 (2021).
- El Agha, E., et al. Mesenchymal stem cells in fibrotic disease. Cell Stem Cell. 21 (2), 166-177 (2017).
- Joe, A. W., et al. Muscle injury activates resident fibro/adipogenic progenitors that facilitate myogenesis. Nature Cell Biology. 12 (2), 153-163 (2010).
- Liu, W., Liu, Y., Lai, X., Kuang, S. Intramuscular adipose is derived from a non-Pax3 lineage and required for efficient regeneration of skeletal muscles. Developmental Biology. 361 (1), 27-38 (2012).
- Scott, R. W., Arostegui, M., Schweitzer, R., Rossi, F. M. V., Underhill, T. M. Hic1 defines quiescent mesenchymal progenitor subpopulations with distinct functions and fates in skeletal muscle regeneration. Cell Stem Cell. 25 (6), 797-813 (2019).
- Uezumi, A., et al. Fibrosis and adipogenesis originate from a common mesenchymal progenitor in skeletal muscle. Journal of Cell Science. 124, 3654-3664 (2011).
- Hogarth, M. W., et al. Fibroadipogenic progenitors are responsible for muscle loss in limb girdle muscular dystrophy 2B. Nature Communications. 10 (1), 2430 (2019).
- Uezumi, A., Fukada, S., Yamamoto, N., Takeda, S., Tsuchida, K. Mesenchymal progenitors distinct from satellite cells contribute to ectopic fat cell formation in skeletal muscle. Nature Cell Biology. 12 (2), 143-152 (2010).
- Stumm, J., et al. Odd skipped-related 1 (Osr1) identifies muscle-interstitial fibro-adipogenic progenitors (FAPs) activated by acute injury. Stem Cell Research. 32, 8-16 (2018).
- Kopinke, D., Roberson, E. C., Reiter, J. F. Ciliary Hedgehog signaling restricts injury-induced adipogenesis. Cell. 170 (2), 340-351 (2017).
- Huang, Y., Das, A. K., Yang, Q. Y., Zhu, M. J., Du, M. Zfp423 promotes adipogenic differentiation of bovine stromal vascular cells. PLoS One. 7 (10), 47496 (2012).
- Sun, Y. -. M., et al. PDGFRα regulated by miR-34a and FoxO1 promotes adipogenesis in porcine intramuscular preadipocytes through Erk signaling pathway. International Journal of Molecular Sciences. 18 (11), 2424 (2017).
- Te, L. J. I., Doherty, C., Correa, J., Batt, J. Identification, isolation, and characterization of fibro-adipogenic progenitors (FAPs) and myogenic progenitors (MPs) in skeletal muscle in the rat. Journal of Visualized Experiments: JoVE. (172), e61750 (2021).
- Lukjanenko, L., et al. Aging disrupts muscle stem cell function by impairing matricellular WISP1 secretion from fibro-adipogenic progenitors. Cell Stem Cell. 24 (3), 433-446 (2019).
- Santini, M. P., et al. Tissue-resident PDGFRalpha(+) progenitor cells contribute to fibrosis versus healing in a context- and spatiotemporally dependent manner. Cell Reports. 30 (2), 555-570 (2020).
- Uezumi, A., et al. Identification and characterization of PDGFRalpha+ mesenchymal progenitors in human skeletal muscle. Cell Death & Disease. 5, 1186 (2014).
- Wosczyna, M. N., et al. Mesenchymal stromal cells are required for regeneration and homeostatic maintenance of skeletal muscle. Cell Reports. 27 (7), 2029-2035 (2019).
- Soliman, H., et al. Pathogenic potential of Hic1-expressing cardiac stromal progenitors. Cell Stem Cell. 26 (2), 205-220 (2020).
- Chung, M. I., Bujnis, M., Barkauskas, C. E., Kobayashi, Y., Hogan, B. L. M. Niche-mediated BMP/SMAD signaling regulates lung alveolar stem cell proliferation and differentiation. Development. 145 (9), (2018).
- Hilgendorf, K. I., et al. Omega-3 fatty acids activate ciliary FFAR4 to control adipogenesis. Cell. 179 (6), 1289-1305 (2019).
- Waisman, A., Norris, A. M., Elías Costa, M., Kopinke, D. Automatic and unbiased segmentation and quantification of myofibers in skeletal muscle. Scientific Reports. 11 (1), 11793 (2021).
- Kang, S. H., Fukaya, M., Yang, J. K., Rothstein, J. D., Bergles, D. E. NG2+ CNS glial progenitors remain committed to the oligodendrocyte lineage in postnatal life and following neurodegeneration. Neuron. 68 (4), 668-681 (2010).
- Srinivas, S., et al. Cre reporter strains produced by targeted insertion of EYFP and ECFP into the ROSA26 locus. BMC Developmental Biology. 1, 4 (2001).
- Lukjanenko, L., Brachat, S., Pierrel, E., Lach-Trifilieff, E., Feige, J. N. Genomic profiling reveals that transient adipogenic activation is a hallmark of mouse models of skeletal muscle regeneration. PLoS One. 8 (8), 71084 (2013).
- Mahdy, M. A., Lei, H. Y., Wakamatsu, J., Hosaka, Y. Z., Nishimura, T. Comparative study of muscle regeneration following cardiotoxin and glycerol injury. Annals of Anatomy = Anatomischer Anzeiger: Official Organ of the Anatomische Gesellscaft. 202, 18-27 (2015).
- Pisani, D. F., Bottema, C. D., Butori, C., Dani, C., Dechesne, C. A. Mouse model of skeletal muscle adiposity: a glycerol treatment approach. Biochemical and Biophysical Research Communications. 396 (3), 767-773 (2010).
- Kawai, H., et al. Experimental glycerol myopathy: a histological study. Acta Neuropathologica. 80 (2), 192-197 (1990).
- Schmittgen, T. D., Livak, K. J. Analyzing real-time PCR data by the comparative CT method. Nature Protocols. 3 (6), 1101-1108 (2008).
- Uezumi, A., et al. Mesenchymal Bmp3b expression maintains skeletal muscle integrity and decreases in age-related sarcopenia. The Journal of Clinical Investigation. 131 (1), 139617 (2021).
- Biferali, B., et al. Prdm16-mediated H3K9 methylation controls fibro-adipogenic progenitors identity during skeletal muscle repair. Science Advances. 7 (23), 9371 (2021).
- Biltz, N. K., Meyer, G. A. A novel method for the quantification of fatty infiltration in skeletal muscle. Skeletal Muscle. 7 (1), (2017).
- Vieites-Prado, A., Renier, N. Tissue clearing and 3D imaging in developmental biology. Development. 148 (18), (2021).
- Gómez-Gaviro, M. V., Sanderson, D., Ripoll, J., Desco, M. Biomedical applications of tissue clearing and three-dimensional imaging in health and disease. iScience. 23 (8), 101432 (2020).
- Ueda, H. R., et al. Tissue clearing and its applications in neuroscience. Nature Reviews Neuroscience. 21 (2), 61-79 (2020).
