Um Guia para Examinar a Formação de Gordura Intramuscular e sua Origem Celular no Músculo Esquelético
Summary
A substituição do tecido muscular saudável por gordura intramuscular é uma característica proeminente das doenças e condições humanas. Este protocolo descreve como visualizar, imagem e quantificar a gordura intramuscular, permitindo o estudo rigoroso dos mecanismos subjacentes à formação de gordura intramuscular.
Abstract
Progenitores fibro-adipogênicos (FAPs) são células mesenquimais estromicas que desempenham um papel crucial durante a homeostase e regeneração muscular esquelética. As FAPs constroem e mantêm a matriz extracelular que age como um andaime molecular de miofibra. Além disso, os FAPs são indispensáveis para a regeneração da miofibra, pois secretam uma infinidade de fatores benéficos sentidos pelas células-tronco musculares (MuSCs). Em estados doentes, no entanto, as FAPs são a origem celular da gordura intramuscular e do tecido cicatricial. Esta fibrose gordurosa é uma marca registrada de sarcopenia e doenças neuromusculares, como distrofia muscular de Duchenne. Uma barreira significativa na determinação do porquê e como as FAPs se diferenciam em gordura intramuscular é a preservação efetiva e posterior visualização de adipócitos, especialmente em seções de tecido congelado. Métodos convencionais de processamento do tecido muscular esquelético, como o congelamento de snaps, não preservam adequadamente a morfologia dos adipócitos individuais, impedindo assim a visualização e quantificação precisas. Para superar esse obstáculo, foi desenvolvido um protocolo rigoroso que preserva a morfologia adipócito em seções musculares esqueléticas que permitem visualização, imagem e quantificação de gordura intramuscular. O protocolo também descreve como processar uma porção de tecido muscular para RT-qPCR, permitindo que os usuários confirmem alterações observadas na formação de gordura, visualizando diferenças na expressão de genes adipogênicos. Além disso, pode ser adaptado para visualizar adipócitos por imunofluorescência de montagem total de amostras musculares. Finalmente, este protocolo descreve como realizar o rastreamento de linhagem genética de FAPs que expressam Pdgfrα para estudar a conversão adipogênica de FAPs. Este protocolo produz consistentemente imagens imunofluorescentes de alta resolução e morfologicamente precisas de adipócitos, juntamente com a confirmação pelo RT-qPCR, permitindo uma visualização e quantificação robusta, rigorosa e reprodutível de gordura intramuscular. Juntos, o pipeline de análise descrito aqui é o primeiro passo para melhorar nossa compreensão de como as FAPs se diferenciam em gordura intramuscular, e fornece uma estrutura para validar novas intervenções para prevenir a formação de gordura.
Introduction
A infiltração de tecido muscular saudável com fibrose gordurosa é uma característica proeminente da Distrofia Muscular de Duchenne (DMD) e outras doenças neuromusculares, bem como sarcopenia, obesidade e diabetes 1,2,3,4,5,6,7,8,9,10 . Embora o aumento da infiltração de gordura nessas condições esteja fortemente associado à diminuição da função muscular, nosso conhecimento sobre o porquê e como as formas de gordura intramuscular ainda são limitadas. Os FAPs são uma população multipotente de células estromal mesenquimais presentes na maioria dos órgãos adultos, incluindo músculo esquelético11,12. Com a idade e as doenças crônicas, no entanto, as FAPs produzem tecido cicatricial fibroso e diferenciam-se em adipócitos, que estão localizados entre os miofibers individuais e formam gordura intramuscular 13,14,15,16,17,18,19,20.
Para começar a combater a formação de gordura intramuscular, os mecanismos de como as FAPs se transformam em adipócitos precisam ser definidos. PDGFRα é o marcador “padrão-ouro” no campo para identificar FAPs dentro do músculo de múltiplas espécies 13,16,17,18,20,21,22,23,24,25,26,27. Como resultado, várias linhas cre indutíveis de tamoxifeno murine, sob o controle do promotor Pdgfrα, foram geradas, permitindo a manipulação genética de FAPs in vivo usando o sistema Cre-LoxP 27,28,29. Por exemplo, combinando esta linha de Cre indutível com um repórter genético, o rastreamento de linhagem de FAPs pode ser realizado, uma estratégia que aplicamos com sucesso ao mapa do destino FAPs em tecido adiposo muscular e branco20,30. Além do rastreamento de linhagem, essas linhas Cre fornecem ferramentas valiosas para estudar a conversão FAP-para-gordura.
Um grande obstáculo na definição do mecanismo da conversão adipogênica das FAPs em gordura intramuscular é a capacidade de quantificar rigorosamente e reprodutivelmente a quantidade de gordura intramuscular que se formou em diferentes condições. A chave é equilibrar a preservação do tecido muscular e adiposo e combinar isso com os métodos de coloração disponíveis para visualizar adipócitos. Por exemplo, o músculo esquelético é muitas vezes congelado sem fixação prévia, preservando miofibers, mas interrompendo a morfologia adipócito (Figura 1). Em contraste, a fixação seguida de incorporação de parafina, enquanto exibe a melhor histologia tecidual, incluindo adipócitos, remove todos os lipídios, tornando assim a maioria dos corantes lipofílicos, como o corante comumente usado Oil Red O, inutilizável.
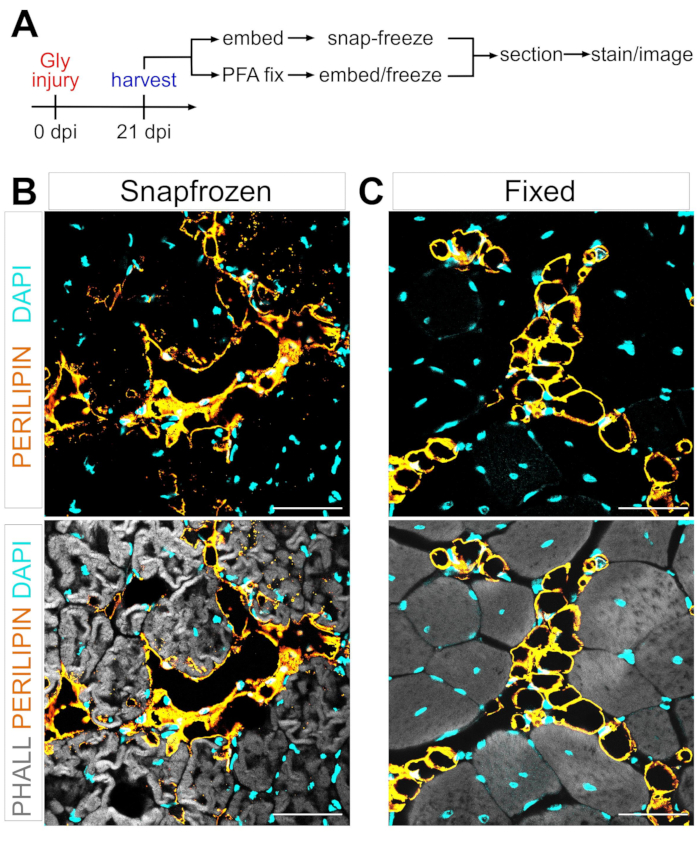
Figura 1: Imagens representativas de gordura intramuscular em tecidos musculares congelados e fixos. (A) Visão geral esquemática da configuração experimental. Imagens imunofluorescentes que mostram adipócitos (amarelo), miofibers (cinza) e núcleos (ciano) dentro de ambos (B) snap-frozen e (C) fixaram TAs em 21 dias após lesão glicerol. Barras de escala: 50 μm. Clique aqui para ver uma versão maior desta figura.
O protocolo aqui descrito preserva a morfologia da miofibra e adipócito e permite a visualização e análise de múltiplos tipos de células. Esta abordagem baseia-se na coloração de imunofluorescência de adipócitos em tecido muscular fixo paraformaldeído (PFA), que permite a co-coloração com múltiplos anticorpos. Também pode ser facilmente adaptado para exibir espacialmente gordura intramuscular em tecido intacto usando imagens de montagem total, fornecendo assim informações sobre o microambiente celular de gordura dentro do músculo. Além disso, este protocolo pode ser combinado com nossa abordagem recentemente publicada para determinar a área transversal de miofibers em tecidos musculares fixos31, uma medida importante para avaliar a saúde muscular. Combinando essa abordagem com o rastreamento de linhagem genética ao mapa do destino, a diferenciação das FAPs em adipócitos também é descrita aqui. Assim, o protocolo versátil aqui descrito permite uma avaliação rigorosa e reprodutível das FAPs e sua diferenciação em gordura intramuscular em seções teciduais e tecidos intactos.
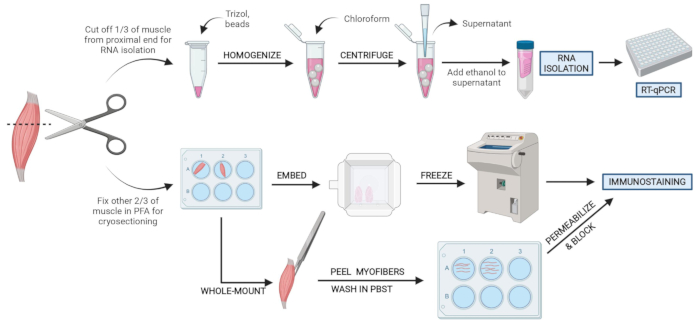
Figura 2: Visão geral do protocolo esquemático. Visão geral esquemática do processamento de tecidos em que um terço do TA é removido, congelado por snap e homogeneizado para análise subsequente de isolamento e transcrição do RNA via RT-qPCR. Os outros dois terços do TA são fixos e processados por PFA para imunossuagem em seções congeladas ou fibras de montagem inteira. Clique aqui para ver uma versão maior desta figura.
Protocol
Representative Results
Discussion
Este protocolo descreve um protocolo extenso e detalhado que permite uma visualização eficiente e uma quantificação rigorosa da gordura intramuscular. Ao dividir o mesmo músculo em duas partes, sendo uma sendo usada para imunofluorescência e outra para análise rt-qPCR, este protocolo também é muito versátil. Também pode ser combinado com o rastreamento de linhagem genética de FAPs para estudar sua conversão em adipócitos sob certas condições e é altamente adaptável ao rótulo e imagem de vários tipos de células adicionais.
As formas mais utilizadas para visualizar a gordura intramuscular são seções de parafina seguidas de manchas de hematoxilina e eosina ou seções congeladas manchadas para corantes lipofílicos, como o Óleo Vermelho O (ORO). No entanto, enquanto os tecidos processados por parafina mantêm a melhor histologia, o mesmo processo também extrai todos os lipídios impedindo o uso de corantes lipofílicos. Embora os métodos de coloração lipofílica funcionem em seções de tecidos fixos e nãofixados da PFA, as gotículas lipídicas são facilmente deslocadas aplicando pressão ao deslizamento de cobertura, distorcendo assim a distribuição espacial da gordura intramuscular. Para contornar isso, um estudo recente estabeleceu um protocolo rigoroso para visualizar adipócitos ORO+ usando uma abordagem de montagem inteira. Para isso, os autores descelularizaram o TA para visualizar a distribuição espacial da gordura intramuscular em toda a TA41. Por mais poderosa que essa técnica seja, também impede o uso de outras co-manchas para marcar estruturas celulares adicionais. Toda a abordagem de imunofluorescência de montagem apresentada aqui pode ser usada para co-manchar adipócitos com uma variedade de marcadores que permitem um mapeamento fino do ambiente celular. Um grande desafio, no entanto, é a penetração tecidual dos anticorpos. Quanto mais fibras forem mantidas juntas, mais difícil será para os anticorpos penetrarem igualmente e amarrarem todos os antígenos disponíveis. Assim, este método é mais eficaz quando se olha para pequenos grupos de fibras. Ao mesmo tempo, esta também é uma limitação, pois a localização anatômica geral da gordura intramuscular está sendo perdida quando se concentra apenas em pequenos feixes de fibras descascados. No entanto, com o desenvolvimento atual de novos métodos de limpeza de tecidos mais novas tecnologias de imagem, maior penetração e visualização de tecidos serão possíveis no futuro 42,43,44.
Embora a fixação prévia do tecido muscular preserve a morfologia adipócito, também cria um desafio para avaliar o tamanho dos miofibers, uma medida importante da saúde muscular. O tamanho da miofibra é determinado medindo a área transversal dos miofibers. Já noticiamos anteriormente que a fixação prévia do tecido muscular fará com que a maioria dos marcadores disponíveis para delinear myofibersfalhem 31. Para superar esse obstáculo, desenvolvemos um novo pipeline de segmentação de imagens, que permite a medição do tamanho da miofibra mesmo nas seções musculares fixas31. Assim, estabelecemos um robusto e eficiente pipeline de processamento de tecidos que, combinado com este protocolo, supera a maioria das desvantagens causadas pela fixação prévia do tecido muscular.
Outra grande vantagem dessa abordagem é a versatilidade. Ao dividir o TA em duas partes, a quantidade de informações que podem ser obtidas de um músculo é maximizada. Isso não só reduz o número de animais, mas também adiciona uma camada extra de controle, confirmando a histologia através da expressão genética e vice-versa. Além disso, muitos genes diferentes podem ser examinados além de genes adipogênicos. O RNA isolado também pode ser usado para um experimento rnaseq muscular inteiro. Finalmente, a peça muscular congelada também pode ser usada para o trabalho proteico. Uma limitação deste protocolo é a possibilidade de a lesão não ser consistente em toda a extensão do TA. Isso pode levar a um cenário em que as duas partes musculares divergem na quantidade de gordura intramuscular que contêm e podem justificar a exclusão de tal amostra de qualquer análise a jusante. Recomenda-se, portanto, não simplesmente confiar no RT-qPCR para tirar grandes conclusões sobre a quantidade de gordura intramuscular, mas sim como dados de apoio às quantificações histológicas.
Em conjunto, este protocolo descreve um robusto, eficiente e rigoroso pipeline de processamento de tecidos que permitirá a visualização e quantificação da gordura intramuscular, o primeiro passo no desenvolvimento de novas opções de tratamento para combater a fibrose gordurosa. Ao mesmo tempo, é versátil e pode ser adaptado a muitos tipos de células diferentes dentro do músculo, bem como adipócitos em outros tecidos.
Disclosures
The authors have nothing to disclose.
Acknowledgements
Agradecemos aos membros do laboratório Kopinke por ajudarem na coleta de dados e na leitura crítica do manuscrito. Agradecemos também aos membros do Instituto de Myology da Universidade da Flórida por sua valiosa contribuição sobre o manuscrito. O trabalho foi apoiado pela subvenção do NIH 1R01AR079449. A Figura 2 foi criada com o Biorender.
Materials
| 16% PFA (Pack of 12, 10 mL bottles) | Electron Miscroscopy Sciences | 15710 | |
| 2.0 mL Microcentrifuge Tubes | Fisher Scientific | 05-408-138 | microcentrifuge tubes for snapfreezing/bead beating |
| 2-Methylbutane (4 L) | Fisher Chemical | O3551-4 | isopentane |
| Absolute Ethanol (200 proof) | ThermoFisher Scientific | BP2818100 | |
| AffiniPure Fab fragment donkey anti-mouse | Jackson ImmunoResearch | 715-007-003 | mouse-on-mouse blocking |
| Alexa Fluor 488 donkey anti-chicken secondary antibody | Jackson ImmunoResearch | 703-545-155 | |
| Alexa Fluor 488 donkey anti-mouse secondary antibody | Invitrogen | A21202 | |
| Alexa Fluor 488 donkey anti-rabbit secondary antibody | Invitrogen | A21206 | |
| Alexa Fluor 568 donkey anti-goat secondary antibody | Invitrogen | A11057 | |
| Alexa Fluor 568 donkey anti-rabbit secondary antibody | Invitrogen | A11037 | |
| Alexa Fluor 568 Phalloidin antibody | Invitrogen | A12380 | Dissolved in 1.5 mL methanol (~66 µM working solution) |
| BioLite 24-well Multidishes | ThermoFisher Scientific | 930186 | 24 well plate for PFA tissue incubation |
| Biometra TOne | analytikjena | 8462070301 | Thermal cycler |
| Chicken anti-GFP antibody | Aves Labs | GFP-1020 | |
| Chloroform/isoamyl alcohol 24:1(v/v) for molecular biology, DNAse, RNAse, and Protease free | ThermoFisher Scientific | AC327155000 | |
| Corn oil | Sigma Aldrich | C8267 | |
| DAPI stain | Invitrogen | D1306 | 150 µM working solution in dH2O |
| Donkey Serum (100 mL) | Millipore Sigma | 5058837 | for blocking solution |
| Dumont #5 Forceps | Fine Science Tools | 11251-20 | sharp-tipped tweezers |
| Fine Scissors Straight 9 cm | Fine Science Tools | 14060-09 | |
| Fluoromount-G | SouthernBiotech | 0100-01 | mounting medium |
| Glycerol, 99.5%, for molecular biology (500 mL) | Acros Organics | 327255000 | |
| Goat anti-PDGFRα antibody | R&D | AF1062 | |
| Hybridization Oven | VWR | 230301V | for Tamoxifen incubation |
| ImmEdge Hydrophobic Barrier PAP Pen | Vector Laboratories | H-4000 | hydrophobic pen |
| Insulin Syringe with Micro-Fine IV needle (28 G) | BD | 329461 | |
| Insulin Syringe with Slip Tip, 1 mL | BD | 329654 | Insulin syringe without needle, for oral gavaging |
| iScript cDNA Synthesis Kit | Bio-Rad | 1708890 | |
| Isoflurane | Patterson Veterinary | 78938441 | |
| Leica DMi8 inverted microscope | Leica | micrscope used for widefield IF and confocal imaging | |
| Micro Slides | VWR | 48311-703 | positively charged microscope slides |
| mouse anti-MYOD antibody | Invitrogen | MA1-41017 | |
| mouse anti-PAX7 antibody (supernatant) | DSHB | AB 428528 | |
| MX35 Premier+ Microtome blades | ThermoFisher Scientific | 3052835 | microtome blades |
| NanoDrop 2000 Spectrophotometer | ThermoFisher Scientific | ND2000 | spectrophotometer for RNA yield |
| Play-Doh | Hasbro | modeling compound | |
| PowerUp SYBR Green Master Mix | ThermoFisher Scientific | A25742 | green dye PCR master mix |
| Puralube Vet Ointment | Puralube | 17033-211-38 | vet ophthalmic ointment |
| QuantStudi 6 Flex Real-Time 384-well PCR System | Applied Biosystems | 4485694 | qPCR machine |
| Rabbit anti-perilipin antibody | Cell Signaling Technology | 9349S | |
| Red-Rotor Shaker | Hoefer Scientific | PR70-115V | shaker for IF staining |
| Richard-Allan Scientific Slip-Rite Cover Glass | ThermoFisher Scientific | 152460 | coverslips |
| RNeasy Mini Kit | QIAGEN | 74106 | contains mini spin columns |
| Safe-Lock Tubes 1.5 ml, natural | Eppendorf | 22363204 | |
| Sample Tubes RB (2 mL) | QIAGEN | 990381 | |
| Sodium azide | Alfa Aesar | 14314 | |
| Stainless Steel Beads, 2.8 mm | Precellys | KT03961-1-101.BK | small beads |
| Stainless Steel Beads, 5 mm | QIAGEN | 69989 | medium beads |
| Stainless Steel Beads, 7 mm | QIAGEN | 69990 | large beads |
| Stainless Steel Disposable Scalpels | Miltex | 327-4102 | scalpel |
| Stainless steel feeding tube, 20 G x 38 mm, straight | Instech Laboratories | FTSS-20S-3 | gavage needle |
| Tamoxifen | Toronto Research Chemicals | T006000 | |
| Tissue Plus O.C.T. Compound | Fisher HealthCare | 4585 | embedding medium |
| TissueLyser LT | QIAGEN | 85600 | bead beater |
| TissueLyser LT Adapter, 12-Tube | QIAGEN | 69980 | |
| Tissue-Tek Cryomold | Sakura | 4566 | specimen molds |
| Triton X-100 | Alfa Aesar | A16046 | |
| TRIzol Reagent | ThermoFisher Scientific | 15596026 | guanidium thiocyanate |
| Tween20 (500 mL) | Fisher BioReagents | BP337-500 | |
| VWR Micro Slides – Superfrost Plus | VWR | 48311703 | |
| Wheaton Coplin staining jars | Millipore Sigma | S6016 | Coplin jar |
References
- Milad, N., et al. Increased plasma lipid levels exacerbate muscle pathology in the mdx mouse model of Duchenne muscular dystrophy. Skeletal Muscle. 7 (1), 19 (2017).
- Goodpaster, B. H., et al. Obesity, regional body fat distribution, and the metabolic syndrome in older men and women. Archives of Internal Medicine. 165 (7), 777-783 (2005).
- Goodpaster, B. H., et al. Association between regional adipose tissue distribution and both type 2 diabetes and impaired glucose tolerance in elderly men and women. Diabetes Care. 26 (2), 372-379 (2003).
- Goodpaster, B. H., et al. The loss of skeletal muscle strength, mass, and quality in older adults: the health, aging and body composition study. The Journals of Gerontology, Series A: Biological Sciences and Medical Sciences. 61 (10), 1059-1064 (2006).
- Goodpaster, B. H., Thaete, F. L., Kelley, D. E. Thigh adipose tissue distribution is associated with insulin resistance in obesity and in type 2 diabetes mellitus. American Journal of Clinical Nutrition. 71 (4), 885-892 (2000).
- Goodpaster, B. H., Theriault, R., Watkins, S. C., Kelley, D. E. Intramuscular lipid content is increased in obesity and decreased by weight loss. Metabolism. 49 (4), 467-472 (2000).
- Burakiewicz, J., et al. Quantifying fat replacement of muscle by quantitative MRI in muscular dystrophy. Journal of Neurology. 264 (10), 2053-2067 (2017).
- Murphy, W. A., Totty, W. G., Carroll, J. E. MRI of normal and pathologic skeletal muscle. American Journal of Roentgenology. 146 (3), 565-574 (1986).
- Willcocks, R. J., et al. Multicenter prospective longitudinal study of magnetic resonance biomarkers in a large duchenne muscular dystrophy cohort. Annals of Neurology. 79 (4), 535-547 (2016).
- Wokke, B. H., et al. Quantitative MRI and strength measurements in the assessment of muscle quality in Duchenne muscular dystrophy. Neuromuscular Disorders. 24 (5), 409-416 (2014).
- Contreras, O., Rossi, F. M. V., Theret, M. Origins, potency, and heterogeneity of skeletal muscle fibro-adipogenic progenitors-time for new definitions. Skeletal Muscle. 11 (1), 16 (2021).
- El Agha, E., et al. Mesenchymal stem cells in fibrotic disease. Cell Stem Cell. 21 (2), 166-177 (2017).
- Joe, A. W., et al. Muscle injury activates resident fibro/adipogenic progenitors that facilitate myogenesis. Nature Cell Biology. 12 (2), 153-163 (2010).
- Liu, W., Liu, Y., Lai, X., Kuang, S. Intramuscular adipose is derived from a non-Pax3 lineage and required for efficient regeneration of skeletal muscles. Developmental Biology. 361 (1), 27-38 (2012).
- Scott, R. W., Arostegui, M., Schweitzer, R., Rossi, F. M. V., Underhill, T. M. Hic1 defines quiescent mesenchymal progenitor subpopulations with distinct functions and fates in skeletal muscle regeneration. Cell Stem Cell. 25 (6), 797-813 (2019).
- Uezumi, A., et al. Fibrosis and adipogenesis originate from a common mesenchymal progenitor in skeletal muscle. Journal of Cell Science. 124, 3654-3664 (2011).
- Hogarth, M. W., et al. Fibroadipogenic progenitors are responsible for muscle loss in limb girdle muscular dystrophy 2B. Nature Communications. 10 (1), 2430 (2019).
- Uezumi, A., Fukada, S., Yamamoto, N., Takeda, S., Tsuchida, K. Mesenchymal progenitors distinct from satellite cells contribute to ectopic fat cell formation in skeletal muscle. Nature Cell Biology. 12 (2), 143-152 (2010).
- Stumm, J., et al. Odd skipped-related 1 (Osr1) identifies muscle-interstitial fibro-adipogenic progenitors (FAPs) activated by acute injury. Stem Cell Research. 32, 8-16 (2018).
- Kopinke, D., Roberson, E. C., Reiter, J. F. Ciliary Hedgehog signaling restricts injury-induced adipogenesis. Cell. 170 (2), 340-351 (2017).
- Huang, Y., Das, A. K., Yang, Q. Y., Zhu, M. J., Du, M. Zfp423 promotes adipogenic differentiation of bovine stromal vascular cells. PLoS One. 7 (10), 47496 (2012).
- Sun, Y. -. M., et al. PDGFRα regulated by miR-34a and FoxO1 promotes adipogenesis in porcine intramuscular preadipocytes through Erk signaling pathway. International Journal of Molecular Sciences. 18 (11), 2424 (2017).
- Te, L. J. I., Doherty, C., Correa, J., Batt, J. Identification, isolation, and characterization of fibro-adipogenic progenitors (FAPs) and myogenic progenitors (MPs) in skeletal muscle in the rat. Journal of Visualized Experiments: JoVE. (172), e61750 (2021).
- Lukjanenko, L., et al. Aging disrupts muscle stem cell function by impairing matricellular WISP1 secretion from fibro-adipogenic progenitors. Cell Stem Cell. 24 (3), 433-446 (2019).
- Santini, M. P., et al. Tissue-resident PDGFRalpha(+) progenitor cells contribute to fibrosis versus healing in a context- and spatiotemporally dependent manner. Cell Reports. 30 (2), 555-570 (2020).
- Uezumi, A., et al. Identification and characterization of PDGFRalpha+ mesenchymal progenitors in human skeletal muscle. Cell Death & Disease. 5, 1186 (2014).
- Wosczyna, M. N., et al. Mesenchymal stromal cells are required for regeneration and homeostatic maintenance of skeletal muscle. Cell Reports. 27 (7), 2029-2035 (2019).
- Soliman, H., et al. Pathogenic potential of Hic1-expressing cardiac stromal progenitors. Cell Stem Cell. 26 (2), 205-220 (2020).
- Chung, M. I., Bujnis, M., Barkauskas, C. E., Kobayashi, Y., Hogan, B. L. M. Niche-mediated BMP/SMAD signaling regulates lung alveolar stem cell proliferation and differentiation. Development. 145 (9), (2018).
- Hilgendorf, K. I., et al. Omega-3 fatty acids activate ciliary FFAR4 to control adipogenesis. Cell. 179 (6), 1289-1305 (2019).
- Waisman, A., Norris, A. M., Elías Costa, M., Kopinke, D. Automatic and unbiased segmentation and quantification of myofibers in skeletal muscle. Scientific Reports. 11 (1), 11793 (2021).
- Kang, S. H., Fukaya, M., Yang, J. K., Rothstein, J. D., Bergles, D. E. NG2+ CNS glial progenitors remain committed to the oligodendrocyte lineage in postnatal life and following neurodegeneration. Neuron. 68 (4), 668-681 (2010).
- Srinivas, S., et al. Cre reporter strains produced by targeted insertion of EYFP and ECFP into the ROSA26 locus. BMC Developmental Biology. 1, 4 (2001).
- Lukjanenko, L., Brachat, S., Pierrel, E., Lach-Trifilieff, E., Feige, J. N. Genomic profiling reveals that transient adipogenic activation is a hallmark of mouse models of skeletal muscle regeneration. PLoS One. 8 (8), 71084 (2013).
- Mahdy, M. A., Lei, H. Y., Wakamatsu, J., Hosaka, Y. Z., Nishimura, T. Comparative study of muscle regeneration following cardiotoxin and glycerol injury. Annals of Anatomy = Anatomischer Anzeiger: Official Organ of the Anatomische Gesellscaft. 202, 18-27 (2015).
- Pisani, D. F., Bottema, C. D., Butori, C., Dani, C., Dechesne, C. A. Mouse model of skeletal muscle adiposity: a glycerol treatment approach. Biochemical and Biophysical Research Communications. 396 (3), 767-773 (2010).
- Kawai, H., et al. Experimental glycerol myopathy: a histological study. Acta Neuropathologica. 80 (2), 192-197 (1990).
- Schmittgen, T. D., Livak, K. J. Analyzing real-time PCR data by the comparative CT method. Nature Protocols. 3 (6), 1101-1108 (2008).
- Uezumi, A., et al. Mesenchymal Bmp3b expression maintains skeletal muscle integrity and decreases in age-related sarcopenia. The Journal of Clinical Investigation. 131 (1), 139617 (2021).
- Biferali, B., et al. Prdm16-mediated H3K9 methylation controls fibro-adipogenic progenitors identity during skeletal muscle repair. Science Advances. 7 (23), 9371 (2021).
- Biltz, N. K., Meyer, G. A. A novel method for the quantification of fatty infiltration in skeletal muscle. Skeletal Muscle. 7 (1), (2017).
- Vieites-Prado, A., Renier, N. Tissue clearing and 3D imaging in developmental biology. Development. 148 (18), (2021).
- Gómez-Gaviro, M. V., Sanderson, D., Ripoll, J., Desco, M. Biomedical applications of tissue clearing and three-dimensional imaging in health and disease. iScience. 23 (8), 101432 (2020).
- Ueda, H. R., et al. Tissue clearing and its applications in neuroscience. Nature Reviews Neuroscience. 21 (2), 61-79 (2020).
