Le domaine de la biologie synthétique ascendante se concentre sur la construction de cellules (minimales) 1,2 et de bioréacteurs métaboliques à des fins biotechnologiques 3,4 ou biomédicales 5,6,7,8. La construction de cellules synthétiques fournit une plate-forme unique qui permet aux chercheurs d’étudier les protéines (membranaires) dans des conditions bien définies imitant celles des environnements natifs, permettant la découverte de propriétés émergentes et de fonctions biochimiques cachées des protéines et des réseaux de réactions9. En tant qu’étape intermédiaire vers une cellule synthétique fonctionnant de manière autonome, des modules sont développés qui capturent les caractéristiques essentielles des cellules vivantes telles que la conservation de l’énergie métabolique, la synthèse des protéines et des lipides et l’homéostasie. De tels modules améliorent non seulement notre compréhension de la vie, mais ont également des applications potentielles dans les domaines de la médecine8 et de la biotechnologie10.
Les protéines transmembranaires sont au cœur de pratiquement tous les réseaux métaboliques, car elles transportent des molécules à l’intérieur ou à l’extérieur de la cellule, signalent et répondent à la qualité de l’environnement et jouent de nombreux rôles biosynthétiques. Ainsi, l’ingénierie de modules métaboliques dans des cellules synthétiques nécessite dans la plupart des cas la reconstitution de protéines membranaires intégrales et/ou périphériques en une bicouche membranaire composée de lipides spécifiques et d’une grande intégrité (faible perméabilité). La manipulation de ces protéines membranaires est un défi et nécessite des connaissances spécifiques et des compétences expérimentales.
Plusieurs méthodes ont été développées pour reconstituer des protéines membranaires au sein des vésicules phospholipidiques, le plus souvent dans le but d’étudier la fonction11,12, la régulation13, les propriétés cinétiques14,15, la dépendance lipidique15,16 et/ou la stabilité17 d’une protéine spécifique. Ces méthodes impliquent la dilution rapide des protéines solubilisées au détergent dans un milieu aqueux en présence de lipides18, l’élimination des détergents par incubation de protéines solubilisées au détergent avec des vésicules lipidiques déstabilisées par le détergent et l’absorption du ou des détergents sur des billes de polystyrène19, ou l’élimination des détergents par dialyse ou chromatographie d’exclusion stérique20. Des solvants organiques ont été utilisés pour former des vésicules lipidiques, par exemple, via la formation d’interphases huile-eau21, mais la majorité des protéines membranaires intégrales sont inactivées lorsqu’elles sont exposées à de tels solvants.
Dans notre laboratoire, nous reconstituons principalement les protéines membranaires par la méthode d’absorption détergente pour former de grandes vésicules unilamellaires (LUVs)19. Cette méthode permet la co-reconstitution de plusieurs protéines membranaires et l’encapsulation dans la lumière vésiculaire d’enzymes, de métabolites et de sondes22,23. Les LUV contenant des protéines membranaires peuvent être convertis en vésicules unilamellaires géantes (GUV) avec/sans encapsulation de composants solubles dans l’eau, en utilisant soit l’électroformation24, soit le gonflement assisté par gel25 et des conditions spécifiques pour préserver l’intégrité des protéines membranaires26.
Cet article présente un protocole pour la reconstitution dans les LUV d’un réseau métabolique hors équilibre qui régénère l’ATP par la décomposition de la L-arginine en L-ornithine27. La formation d’ATP est couplée à la production de glycérol-3-phosphate (G3P), un élément constitutif important pour la synthèse des phospholipides22,28. La voie métabolique se compose de deux protéines membranaires intégrales, une arginine/ornithine (ArcD) et un antiporteur G3P/Pi (GlpT). De plus, trois enzymes solubles (ArcA, ArcB, ArcC) sont nécessaires au recyclage de l’ATP, et GlpK est utilisé pour convertir le glycérol en glycérol 3-phosphate, en utilisant l’ATP issu de la dégradation de la L-arginine, voir la figure 1 pour une vue d’ensemble schématique de la voie. Ce protocole représente un bon point de départ pour la construction future de réseaux réactionnels encore plus complexes, pour la synthèse de lipides ou de protéines ou la division des cellules. La composition lipidique des vésicules soutient l’activité d’une grande variété de protéines membranaires intégrales et a été optimisée pour le transport de diverses molécules dans ou hors des vésicules 27,29,30.
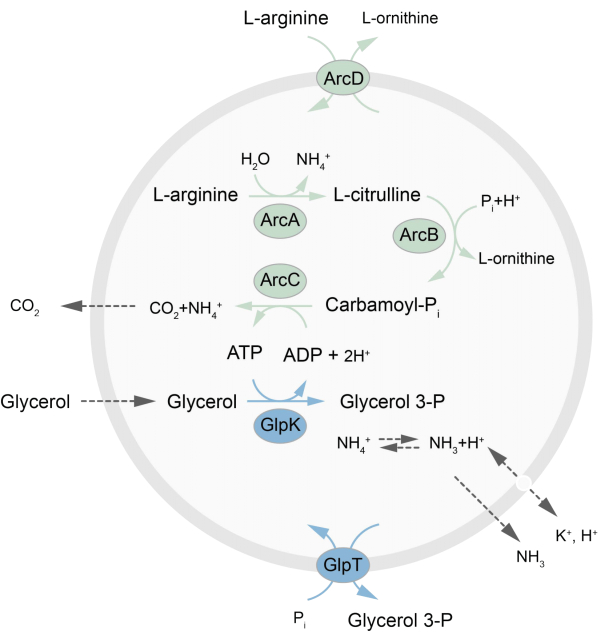
Figure 1 : Vue d’ensemble de la voie de production de l’ATP et de la synthèse et de l’excrétion du glycérol 3-phosphate. Veuillez cliquer ici pour voir une version agrandie de cette figure.
En bref, des protéines membranaires purifiées (solubilisées dans du dodécyl-β-D-maltoside, DDM) sont ajoutées à des vésicules lipidiques préformées qui ont été déstabilisées avec Triton X-100, ce qui permet l’insertion des protéines dans la membrane. Les molécules de détergent sont ensuite (lentement) éliminées par l’ajout de billes de polystyrène activées, ce qui entraîne la formation de protéoliposomes bien scellés. Des composants solubles peuvent ensuite être ajoutés aux vésicules et encapsulés via des cycles de gel-dégel, ce qui piège les molécules dans le processus de fusion membranaire. Les vésicules obtenues sont très hétérogènes et beaucoup sont multilamellaires. Ils sont ensuite extrudés à travers un filtre en polycarbonate avec une taille de pores de 400, 200 ou 100 nm, ce qui donne des vésicules de taille plus uniforme ; Plus la taille des pores est petite, plus les vésicules sont homogènes et unilamellaires, mais au prix d’un volume interne plus petit. Les protéines non incorporées et les petites molécules sont éliminées de la solution externe par chromatographie d’exclusion stérique. Les proteoLUV peuvent être convertis en vésicules de taille micrométrique par gonflement assisté par gel, et ces proteoGUVs sont ensuite collectés et piégés dans une puce microfluidique pour la caractérisation et la manipulation microscopiques. La figure 2 présente une vue d’ensemble schématique de l’ensemble du protocole.
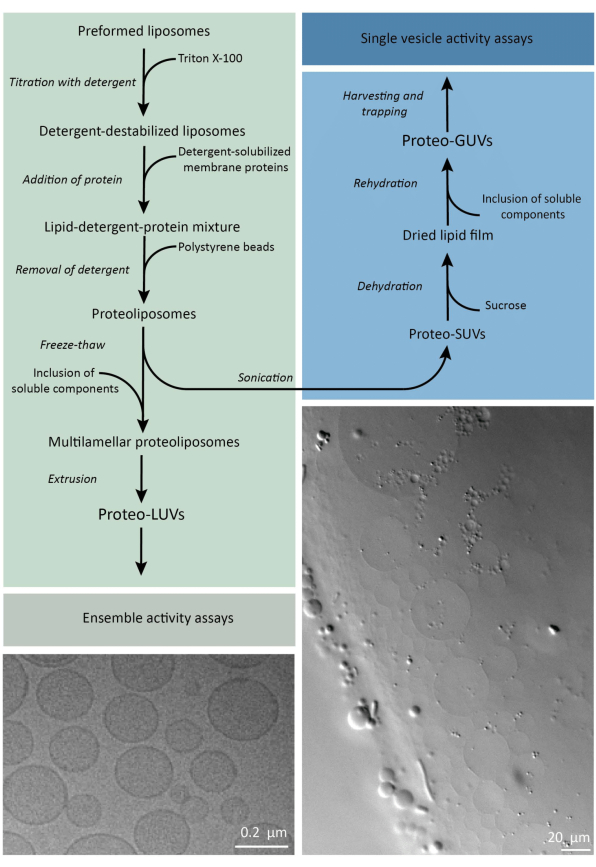
Figure 2 : Vue d’ensemble du protocole de reconstitution des protéines membranaires et d’encapsulation d’enzymes et de composants hydrosolubles dans des vésicules lipidiques submicrométriques (LUV) et micrométriques (GUV). Veuillez cliquer ici pour voir une version agrandie de cette figure.
Les protocoles de reconstitution et d’encapsulation fonctionnent bien et la fonctionnalité des protéines est conservée, mais les proteoLUVs et les proteoGUVs sont de taille hétérogène. Les approches microfluidiques31,32 permettent la formation de vésicules de taille micrométrique de taille plus homogène, mais la reconstitution fonctionnelle des protéines membranaires n’est généralement pas possible car le solvant résiduel dans la bicouche inactive les protéines. La taille des protéoLUV varie de 100 à 400 nm, et à de faibles concentrations d’enzymes, l’encapsulation peut conduire à des vésicules avec des voies métaboliques incomplètes (effets stochastiques ; voir Figure 3). Les LUV sont idéaux pour construire des modules métaboliques spécifiques, comme le montre ici la production d’ATP et de blocs de construction comme le G3P. De tels protéoLUV peuvent potentiellement être encapsulés dans des GUV et servir de compartiments semblables à des organites pour les vésicules hôtes.
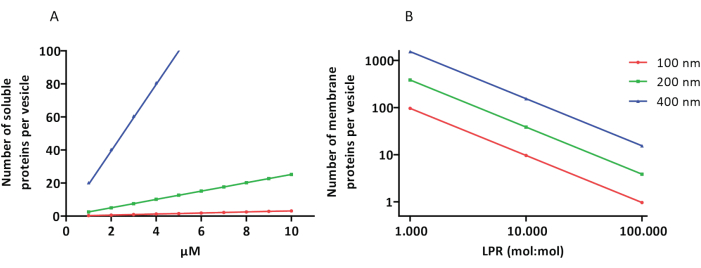
Figure 3 : Nombre de molécules par vésicule d’un diamètre de 100, 200 ou 400 nm. (A) Lorsque les protéines encapsulées (enzymes, sondes) sont dans la gamme de 1 à 10 μM. (B) La reconstitution se fait à 1 à 1 000, 1 à 10 000 et 1 à 100 000 protéines membranaires par lipide (mol/mol). Nous faisons l’hypothèse que les molécules sont encapsulées aux concentrations indiquées et incorporées dans la membrane à ces ratios protéines/lipides. Pour certaines enzymes, nous avons vu qu’elles se lient aux membranes, ce qui peut augmenter leur concentration apparente dans les vésicules. Abréviation : LPR = Lipid-Protein-Ratio Veuillez cliquer ici pour voir une version agrandie de ce chiffre.
