Het vakgebied van bottom-up synthetische biologie richt zich op het bouwen van (minimale) cellen 1,2 en metabole bioreactoren voor biotechnologische 3,4 of biomedische doeleinden 5,6,7,8. De constructie van synthetische cellen biedt een uniek platform dat onderzoekers in staat stelt om (membraan)eiwitten te bestuderen in goed gedefinieerde omstandigheden die die van inheemse omgevingen nabootsen, waardoor opkomende eigenschappen en verborgen biochemische functies van eiwitten en reactienetwerken kunnen worden ontdekt9. Als tussenstap naar een autonoom functionerende synthetische cel worden modules ontwikkeld die essentiële kenmerken van levende cellen vastleggen, zoals metabolisch energiebehoud, eiwit- en lipidensynthese en homeostase. Dergelijke modules vergroten niet alleen ons begrip van het leven, maar hebben ook potentiële toepassingen op het gebied van geneeskunde8 en biotechnologie10.
Transmembraaneiwitten vormen de kern van vrijwel elk metabolisch netwerk, omdat ze moleculen in of uit de cel transporteren, signalen geven en reageren op de kwaliteit van de omgeving, en tal van biosynthetische rollen spelen. De engineering van metabole modules in synthetische cellen vereist dus in de meeste gevallen de reconstitutie van integrale en/of perifere membraaneiwitten tot een membraandubbellaag die is samengesteld uit specifieke lipiden en een hoge integriteit (lage permeabiliteit). De omgang met deze membraaneiwitten is uitdagend en vereist specifieke kennis en experimentele vaardigheden.
Er zijn verschillende methoden ontwikkeld om membraaneiwitten in fosfolipideblaasjes te reconstrueren, meestal met als doel de functie11,12, regulatie13, kinetische eigenschappen14,15, lipideafhankelijkheid15,16 en/of stabiliteit17 van een specifiek eiwit te bestuderen. Deze methoden omvatten de snelle verdunning van detergent-oplosbaar eiwit in waterige media in aanwezigheid van lipiden18, de verwijdering van detergenten door het incuberen van detergent-solubel eiwit met detergent-gedestabiliseerde lipideblaasjes en absorptie van het detergent(en) op polystyreenkorrels19, of het verwijderen van detergenten door dialyse of grootte-uitsluitingschromatografie20. Organische oplosmiddelen zijn gebruikt om lipideblaasjes te vormen, bijvoorbeeld via de vorming van olie-water-interfasen21, maar de meeste integrale membraaneiwitten worden geïnactiveerd wanneer ze worden blootgesteld aan dergelijke oplosmiddelen.
In ons laboratorium reconstrueren we meestal membraaneiwitten door middel van de detergent-absorptiemethode om grote unilamellaire blaasjes (LUV’s) te vormen19. Deze methode maakt de co-reconstitutie van meerdere membraaneiwitten en de inkapseling in het blaasjeslumen van enzymen, metabolieten en sondes mogelijk22,23. De membraaneiwitbevattende LUV’s kunnen worden omgezet in reusachtige-unilamellaire blaasjes (GUV’s) met/zonder inkapseling van in water oplosbare componenten, met behulp van elektroformatie24 of gelondersteunde zwelling25 en specifieke omstandigheden om de integriteit van de membraaneiwitten te behouden26.
Dit artikel presenteert een protocol voor de reconstitutie in LUV’s van een uit evenwicht zijnd metabolisch netwerk dat ATP regenereert door de afbraak van L-arginine in L-ornithine27. De vorming van ATP gaat gepaard met de productie van glycerol-3-fosfaat (G3P), een belangrijke bouwsteen voor de fosfolipidensynthese22,28. De metabole route bestaat uit twee integrale membraaneiwitten, een arginine/ornithine (ArcD) en een G3P/Pi-antiporter (GlpT). Daarnaast zijn drie oplosbare enzymen (ArcA, ArcB, ArcC) nodig voor de recycling van ATP, en GlpK wordt gebruikt om glycerol om te zetten in glycerol 3-fosfaat, waarbij gebruik wordt gemaakt van het ATP uit de afbraak van L-arginine, zie figuur 1 voor een schematisch overzicht van de route. Dit protocol vormt een goed uitgangspunt voor de toekomstige constructie van nog complexere reactienetwerken – voor de synthese van lipiden of eiwitten of de deling van cellen. De lipidensamenstelling van de blaasjes ondersteunt de activiteit van een breed scala aan integrale membraaneiwitten en is geoptimaliseerd voor het transport van diverse moleculen in of uit de blaasjes 27,29,30.
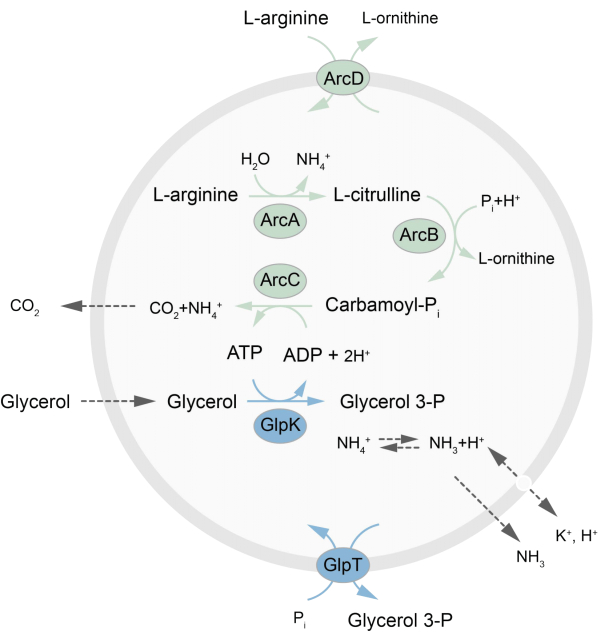
Figuur 1: Overzicht van de route voor ATP-productie en glycerol-3-fosfaatsynthese en -uitscheiding. Klik hier om een grotere versie van deze figuur te bekijken.
Kortom, gezuiverde membraaneiwitten (opgelost in dodecyl-β-D-maltoside, DDM) worden toegevoegd aan voorgevormde lipideblaasjes die zijn gedestabiliseerd met Triton X-100, waardoor de eiwitten in het membraan kunnen worden ingebracht. De wasmiddelmoleculen worden vervolgens (langzaam) verwijderd door de toevoeging van geactiveerde polystyreenkorrels, wat resulteert in de vorming van goed afgesloten proteoliposomen. Oplosbare componenten kunnen vervolgens aan de blaasjes worden toegevoegd en worden ingekapseld via vries-dooicycli, waardoor de moleculen worden opgesloten in het proces van membraanfusie. De verkregen blaasjes zijn zeer heterogeen en veel zijn multilamellair. Ze worden vervolgens geëxtrudeerd door een polycarbonaatfilter met een poriegrootte van 400, 200 of 100 nm, wat blaasjes van meer uniforme grootte oplevert; Hoe kleiner de poriegrootte, hoe homogener en unilamellair de blaasjes, maar tegen de prijs van een kleiner inwendig volume. Niet-opgenomen eiwitten en kleine moleculen worden uit de externe oplossing verwijderd door middel van grootte-uitsluitingschromatografie. De proteoLUV’s kunnen worden omgezet in blaasjes ter grootte van een micrometer door middel van gel-geassisteerde zwelling, en deze proteoGUV’s worden vervolgens verzameld en gevangen in een microfluïdische chip voor microscopische karakterisering en manipulatie. Figuur 2 geeft een schematisch overzicht van het volledige protocol.
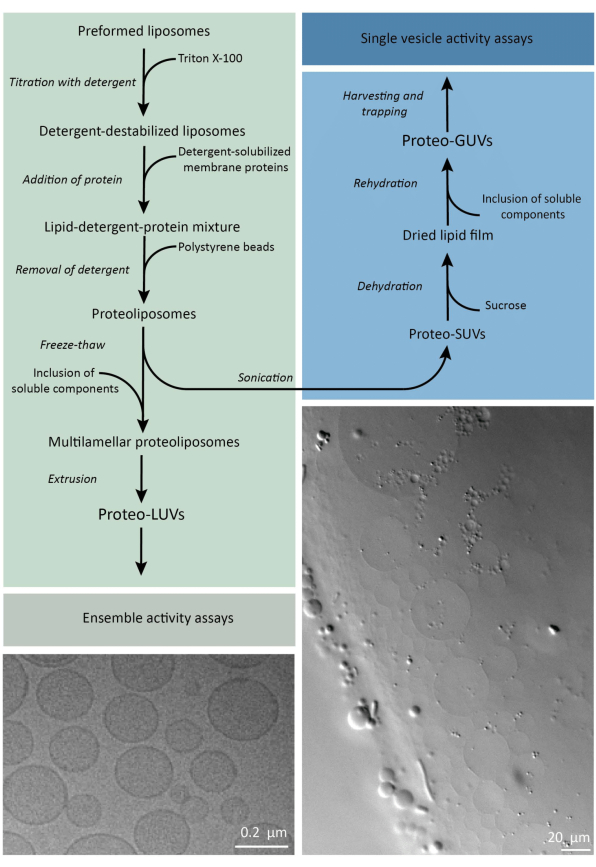
Figuur 2: Overzicht van het protocol voor het reconstitueren van membraaneiwitten en het inkapselen van enzymen en wateroplosbare componenten in lipideblaasjes met een submicrometer (LUV’s) en micrometergrootte (GUV’s). Klik hier om een grotere versie van deze figuur te bekijken.
De reconstitutie- en inkapselingsprotocollen werken goed en de functionaliteit van de eiwitten blijft behouden, maar de proteoLUV’s en proteoGUV’s zijn heterogeen van grootte. Microfluïdische benaderingen31,32 maken de vorming van blaasjes ter grootte van een micrometer mogelijk die homogener van grootte zijn, maar functionele reconstitutie van membraaneiwitten is over het algemeen niet mogelijk omdat resterend oplosmiddel in de dubbellaag de eiwitten inactiveert. De proteoLUV’s variëren in grootte van 100 tot 400 nm, en bij lage concentraties enzymen kan de inkapseling leiden tot blaasjes met onvolledige metabole routes (stochastische effecten; zie figuur 3). LUV’s zijn ideaal voor het bouwen van specifieke metabolische modules, zoals hier te zien is voor de productie van ATP en bouwstenen zoals G3P. Dergelijke proteoLUV’s kunnen mogelijk worden ingekapseld in GUV’s en dienen als organelachtige compartimenten voor de gastheerblaasjes.
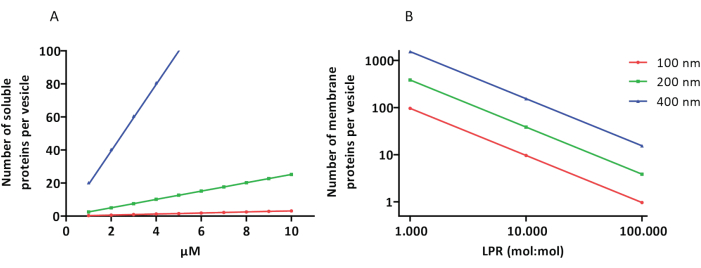
Figuur 3: Aantal moleculen per blaasje met een diameter van 100, 200 of 400 nm. (A) Wanneer de ingekapselde eiwitten (enzymen, sondes) zich in het bereik van 1-10 μM bevinden. (B) De reconstitutie wordt gedaan bij 1 tot 1.000, 1 tot 10.000 en 1 tot 100.000 membraaneiwitten per lipide (mol/mol). We gaan ervan uit dat moleculen in de aangegeven concentraties worden ingekapseld en in het membraan worden opgenomen in deze eiwit-lipideverhoudingen. Voor sommige enzymen hebben we gezien dat ze zich binden aan membranen, waardoor hun schijnbare concentratie in de blaasjes kan toenemen. Afkorting: LPR = Lipid-Protein-Ratio Klik hier om een grotere versie van deze figuur te bekijken.
