Область синтетической биологии снизу вверх фокусируется на конструировании (минимальных) клеток 1,2 и метаболических биореакторов для биотехнологических 3,4 или биомедицинских целей 5,6,7,8. Конструирование синтетических клеток обеспечивает уникальную платформу, которая позволяет исследователям изучать (мембранные) белки в четко определенных условиях, имитирующих условия нативной среды, что позволяет обнаружить эмерджентные свойства и скрытые биохимические функции белков и реакционных сетей9. В качестве промежуточного шага на пути к автономно функционирующей синтетической клетке разрабатываются модули, которые улавливают основные характеристики живых клеток, такие как сохранение метаболической энергии, синтез белков и липидов, а также гомеостаз. Такие модули не только улучшают наше понимание жизни,но и имеют потенциальное применение в области медицины и биотехнологии10.
Трансмембранные белки лежат в основе практически любой метаболической сети, поскольку они транспортируют молекулы внутрь или из клетки, сигнализируют и реагируют на качество окружающей среды, а также играют многочисленные биосинтетические роли. Таким образом, инженерия метаболических модулей в синтетических клетках в большинстве случаев требует восстановления интегральных и/или периферических мембранных белков в мембранный бислой, состоящий из специфических липидов и обладающих высокой целостностью (низкой проницаемостью). Работа с этими мембранными белками сложна и требует специальных знаний и экспериментальных навыков.
Было разработано несколько методов для восстановления мембранных белков в фосфолипидных везикулах, чаще всего с целью изучения функции11,12, регуляции13, кинетических свойств14,15, липидной зависимости 15,16 и/или стабильности17 конкретного белка. Эти методы включают быстрое разведение растворимого в детергентах белка в водную среду в присутствии липидов18, удаление детергентов путем инкубации растворимого в моющем средстве белка с дестабилизированными липидными везикулами и абсорбцию детергента(ов) на гранулах полистирола19 или удаление детергентов с помощью диализа или эксклюзионной хроматографии20. Органические растворители использовались для образования липидных везикул, например, путем образования интерфаз21 нефть-вода, но большинство интегральных мембранных белков инактивируются при воздействии таких растворителей.
В нашей лаборатории мы в основном восстанавливаем мембранные белки методом детергент-абсорбции с образованием крупных униламеллярных везикул (LUV)19. Этот метод позволяет осуществлять совместное воссоздание множественных мембранных белков и инкапсуляцию в просвет везикулы ферментов, метаболитов и зондов22,23. Мембранные белки, содержащие LUV, могут быть преобразованы в гигантские униламеллярные везикулы (GUV) с инкапсуляцией водорастворимых компонентов или без нее, используя либо электрообразование24, либо гелеобразное набухание25 и специальные условия для сохранения целостности мембранных белков26.
В данной статье представлен протокол восстановления в LUV неравновесной метаболической сети, которая регенерирует АТФ путем распада L-аргинина на L-орнитин27. Образование АТФ связано с образованием глицерол-3-фосфата (G3P), важного строительного блока для синтеза фосфолипидов22,28. Метаболический путь состоит из двух интегральных мембранных белков: аргинина/орнитина (ArcD) и антипорта G3P/Pi (GlpT). Кроме того, три растворимых фермента (ArcA, ArcB, ArcC) необходимы для рециркуляции АТФ, а GlpK используется для превращения глицерина в глицерин-3-фосфат с использованием АТФ из распада L-аргинина, см. Рисунок 1 для схематического обзора пути. Этот протокол представляет собой хорошую отправную точку для будущего построения еще более сложных реакционных сетей — для синтеза липидов или белков или для деления клеток. Липидный состав везикул поддерживает активность широкого спектра интегральных мембранных белков и был оптимизирован для транспортировки различных молекул внутрь или из везикул 27,29,30.
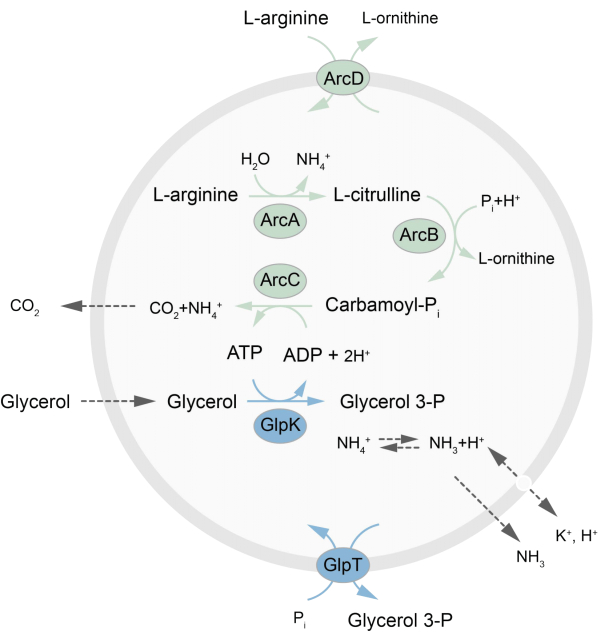
Рисунок 1: Обзор пути производства АТФ, синтеза и выведения глицерин-3-фосфата. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Короче говоря, очищенные мембранные белки (солюбилизированные в додецил-β-D-мальтозиде, ДДМ) добавляются к предварительно сформированным липидным везикулам, которые были дестабилизированы с помощью Triton X-100, что позволяет встраивать белки в мембрану. Молекулы детергента впоследствии (медленно) удаляются путем добавления активированных гранул полистирола, что приводит к образованию хорошо запечатанных протеолипосом. Растворимые компоненты затем могут быть добавлены в везикулы и инкапсулированы с помощью циклов замораживания-оттаивания, что улавливает молекулы в процессе слияния мембран. Полученные везикулы очень неоднородны и многие из них являются многопластинчатыми. Затем они выдавливаются через поликарбонатный фильтр с размером пор 400, 200 или 100 нм, что дает пузырьки более равномерного размера; Чем меньше размер пор, тем более однородными и одноламеллярными являются везикулы, но ценой меньшего внутреннего объема. Неинкорпорированные белки и малые молекулы удаляются из наружного раствора с помощью эксклюзионной хроматографии. ПротеоЛУВ могут быть преобразованы в микрометровые везикулы путем гелеобразного набухания, а затем эти протеоГУВ собираются и удерживаются в микрофлюидном чипе для микроскопической характеристики и манипуляций. На рисунке 2 показан схематический обзор полного протокола.
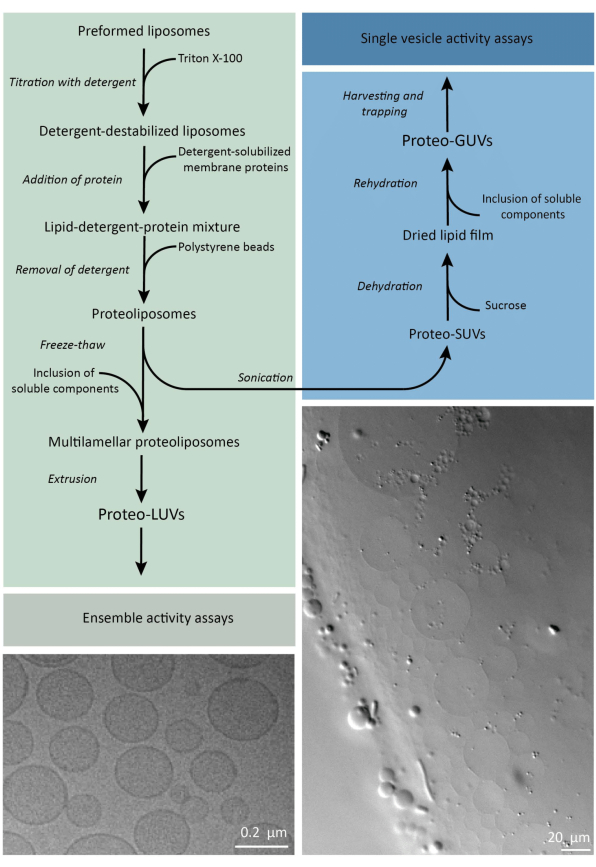
Рисунок 2: Обзор протокола восстановления мембранных белков и инкапсуляции ферментов и водорастворимых компонентов в липидных везикулах субмикронного (LUV) и микрометрового размера (GUVs). Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Протоколы восстановления и инкапсуляции работают хорошо, и функциональность белков сохраняется, но proteoLUV и proteoGUVs неоднородны по размеру. Микрофлюидные подходы31,32 позволяют формировать везикулы микрометрового размера, которые являются более однородными по размеру, но функциональное восстановление мембранных белков, как правило, невозможно, поскольку остаточный растворитель в бислое инактивирует белки. Размер протеоЛУВ колеблется от 100 до 400 нм, и при низких концентрациях ферментов инкапсуляция может привести к образованию везикул с неполными метаболическими путями (стохастические эффекты; см. рис. 3). LUV идеально подходят для создания определенных метаболических модулей, как показано здесь для производства АТФ и строительных блоков, таких как G3P. Такие proteoLUV потенциально могут быть инкапсулированы в GUV и служить органеллоподобными компартментами для везикул хозяина.
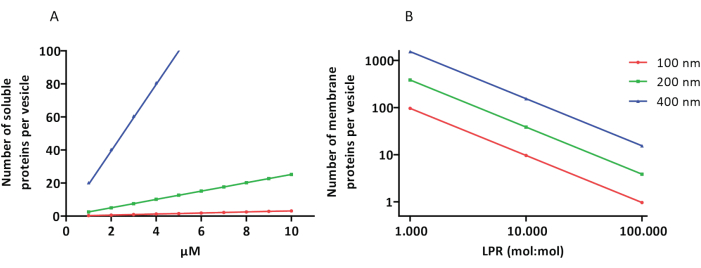
Рисунок 3: Количество молекул в везикуле диаметром 100, 200 или 400 нм. (А) Когда инкапсулированные белки (ферменты, зонды) находятся в диапазоне от 1 до 10 мкМ. (В) Восстановление осуществляется при концентрации от 1 до 1 000, от 1 до 10 000 и от 1 до 100 000 мембранных белков на липид (моль/моль). Мы исходим из предположения, что молекулы инкапсулируются в указанных концентрациях и встраиваются в мембрану при этих соотношении белка к липидам. Что касается некоторых ферментов, то мы видели, что они связываются с мембранами, что может увеличивать их видимую концентрацию в везикулах. Аббревиатура: LPR = Lipid-Protein-Ratio Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
