يركز مجال البيولوجيا التركيبية من أسفل إلى أعلى على بناء الخلايا (الحد الأدنى) 1,2 والمفاعلات الحيوية الأيضية للتكنولوجيا الحيوية 3,4 أو الأغراض الطبية الحيوية 5,6,7,8. يوفر بناء الخلايا الاصطناعية منصة فريدة تسمح للباحثين بدراسة البروتينات (الغشائية) في ظروف محددة جيدا تحاكي تلك الموجودة في البيئات الأصلية ، مما يتيح اكتشاف الخصائص الناشئة والوظائف الكيميائية الحيوية المخفية للبروتينات وشبكات التفاعل9. كخطوة وسيطة نحو خلية اصطناعية تعمل بشكل مستقل ، يتم تطوير وحدات تلتقط السمات الأساسية للخلايا الحية مثل الحفاظ على الطاقة الأيضية ، وتخليق البروتين والدهون ، والتوازن. هذه الوحدات لا تعزز فهمنا للحياة فحسب ، بل لها أيضا تطبيقات محتملة في مجالات الطب8 والتكنولوجيا الحيوية10.
تقع البروتينات عبر الغشاء في قلب أي شبكة أيضية تقريبا لأنها تنقل الجزيئات داخل الخلية أو خارجها ، وتشير ، وتستجيب لجودة البيئة ، وتلعب العديد من أدوار التخليق الحيوي. وبالتالي ، فإن هندسة الوحدات الأيضية في الخلايا الاصطناعية تتطلب في معظم الحالات إعادة تكوين بروتينات الغشاء المتكاملة و / أو المحيطية في طبقة ثنائية غشائية تتكون من دهون محددة وسلامة عالية (نفاذية منخفضة). يمثل التعامل مع هذه البروتينات الغشائية تحديا ويتطلب معرفة محددة ومهارات تجريبية.
تم تطوير عدة طرق لإعادة تكوين البروتينات الغشائية داخل حويصلات الفوسفوليبيد ، غالبا بهدف دراسة الوظيفة11،12 ، والتنظيم13 ، والخصائص الحركية14،15 ، والاعتماد على الدهون15،16 ، و / أو الاستقرار17 لبروتين معين. تتضمن هذه الطرق التخفيف السريع للبروتين القابل للذوبان في المنظفات في وسط مائي في وجود الدهون18 ، وإزالة المنظفات عن طريق احتضان البروتين القابل للذوبان في المنظفات مع حويصلات دهنية غير مستقرة المنظفات وامتصاص المنظف (المنظفات) على حبات البوليسترين19 ، أو إزالة المنظفات عن طريق غسيل الكلى أو كروماتوغرافيا استبعاد الحجم20. تم استخدام المذيبات العضوية لتشكيل حويصلات دهنية ، على سبيل المثال ، عن طريق تكوين الأطوار البينية بين الزيتوالماء 21 ، ولكن غالبية البروتينات الغشائية المتكاملة يتم تعطيلها عند تعرضها لهذه المذيبات.
في مختبرنا ، نقوم في الغالب بإعادة تكوين البروتينات الغشائية بطريقة امتصاص المنظفات لتشكيل حويصلات كبيرة أحادية الصفيحة (LUVs) 19. تسمح هذه الطريقة بإعادة التكوين المشترك لبروتينات غشائية متعددة وتغليف في تجويف الحويصلة للإنزيمات والمستقلبات والمجسات22,23. يمكن تحويل LUVs المحتوية على البروتين الغشائي إلى حويصلات عملاقة أحادية الصفيحة (GUVs) مع / بدون تغليف المكونات القابلة للذوبان في الماء ، إما باستخدام التكوين الكهربائي24 أو التورم بمساعدة الهلام25 وظروف محددة للحفاظ على سلامة بروتينات الغشاء26.
تقدم هذه الورقة بروتوكولا لإعادة تكوين شبكة التمثيل الغذائي خارج التوازن في LUVs التي تجدد ATP من خلال انهيار L-arginine إلى L-ornithine27. يقترن تكوين ATP بإنتاج الجلسرين -3 فوسفات (G3P) ، وهو لبنة بناء مهمة لتخليق الفوسفوليبيد22,28. يتكون المسار الأيضي من بروتينين غشائيين متكاملين ، أرجينين / أورنيثين (ArcD) ومضاد G3P / Pi (GlpT). بالإضافة إلى ذلك ، هناك حاجة إلى ثلاثة إنزيمات قابلة للذوبان (ArcA ، ArcB ، ArcC) لإعادة تدوير ATP ، ويستخدم GlpK لتحويل الجلسرين إلى جلسرين 3-فوسفات ، باستخدام ATP من انهيار L-arginine ، انظر الشكل 1 للحصول على نظرة عامة تخطيطية للمسار. يمثل هذا البروتوكول نقطة انطلاق جيدة للبناء المستقبلي لشبكات تفاعل أكثر تعقيدا – لتخليق الدهون أو البروتينات أو تقسيم الخلايا. يدعم التركيب الدهني للحويصلات نشاط مجموعة واسعة من البروتينات الغشائية المتكاملة وقد تم تحسينه لنقل جزيئات متنوعة داخل أو خارج الحويصلات27،29،30.
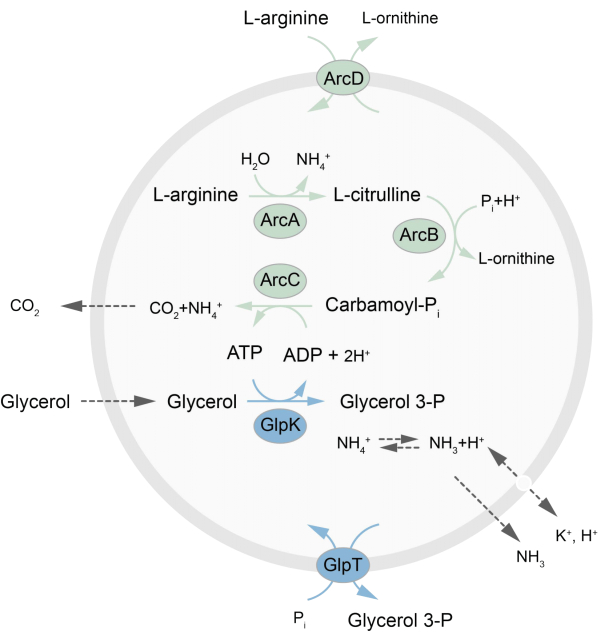
الشكل 1: نظرة عامة على مسار إنتاج الأدينوسين الثلاثي الفوسفات وتخليق وإفراز الجليسرول 3-فوسفات. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
باختصار ، تضاف بروتينات الغشاء المنقى (القابلة للذوبان في دوديسيل β-د-مالتوسيد ، DDM) إلى حويصلات الدهون المشكلة مسبقا والتي تم زعزعة استقرارها باستخدام Triton X-100 ، مما يسمح بإدخال البروتينات في الغشاء. تتم إزالة جزيئات المنظفات لاحقا (ببطء) عن طريق إضافة حبات البوليسترين المنشط ، مما يؤدي إلى تكوين بروتيليبوزومات محكمة الغلق. يمكن بعد ذلك إضافة المكونات القابلة للذوبان إلى الحويصلات وتغليفها عبر دورات التجميد والذوبان ، والتي تحبس الجزيئات في عملية اندماج الغشاء. الحويصلات التي تم الحصول عليها غير متجانسة للغاية والعديد منها متعدد الصفائح. ثم يتم بثقها من خلال مرشح من البولي كربونات بحجم مسام 400 أو 200 أو 100 نانومتر ، مما ينتج عنه حويصلات ذات حجم أكثر اتساقا ؛ كلما كان حجم المسام أصغر ، كانت الحويصلات أكثر تجانسا وأحادية الصفيحة ولكن بسعر حجم داخلي أصغر. تتم إزالة البروتينات غير المدمجة والجزيئات الصغيرة من المحلول الخارجي عن طريق كروماتوغرافيا استبعاد الحجم. يمكن تحويل البروتيوLUVs إلى حويصلات بحجم ميكرومتر عن طريق التورم بمساعدة الهلام ، ثم يتم جمع هذه البروتيو GUVs واحتجازها في رقاقة الموائع الدقيقة للتوصيف المجهري والمعالجة. يوضح الشكل 2 نظرة عامة تخطيطية للبروتوكول الكامل.
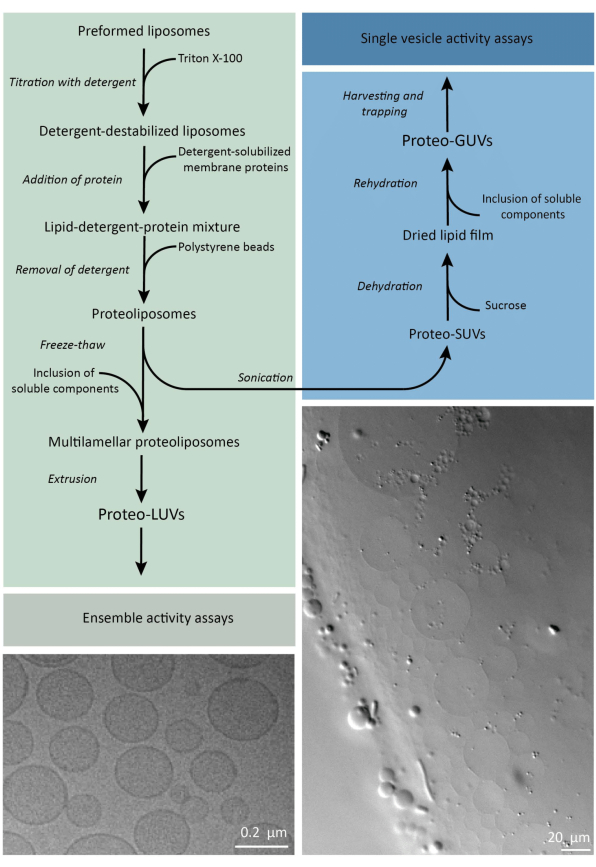
الشكل 2: نظرة عامة على بروتوكول إعادة تكوين البروتينات الغشائية وتغليف الإنزيمات والمكونات القابلة للذوبان في الماء في الحويصلات الدهنية ذات الميكرومتر الفرعي (LUVs) وحجم الميكرومتر (GUVs). يرجى النقر هنا لعرض نسخة أكبر من هذا الشكل.
تعمل بروتوكولات إعادة التكوين والتغليف بشكل جيد ويتم الاحتفاظ بوظائف البروتينات ، لكن البروتيوLUVs و proteoGUVs غير متجانسة في الحجم. تسمح مناهج الموائع الدقيقة31,32 بتكوين حويصلات بحجم ميكرومتر تكون أكثر تجانسا في الحجم ، لكن إعادة التكوين الوظيفي لبروتينات الغشاء غير ممكنة بشكل عام لأن المذيب المتبقي في الطبقة الثنائية يعطل البروتينات. يتراوح حجم البروتيوLUVs من 100 إلى 400 نانومتر ، وعند تركيزات منخفضة من الإنزيمات ، قد يؤدي التغليف إلى حويصلات ذات مسارات أيضية غير مكتملة (التأثيرات العشوائية ؛ انظر الشكل 3). تعد LUVs مثالية لبناء وحدات أيضية محددة ، كما هو موضح هنا لإنتاج ATP وكتل البناء مثل G3P. يمكن تغليف هذه البروتيوLUVs في GUVs وتعمل كمقصورات تشبه العضيات للحويصلات المضيفة.
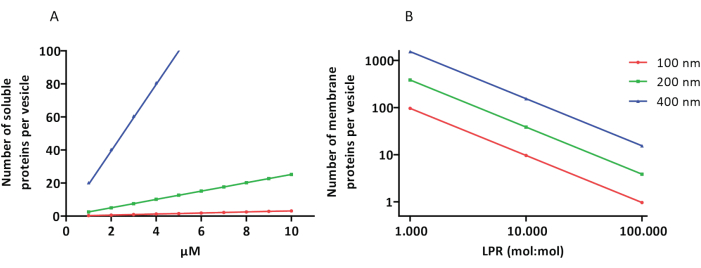
الشكل 3: عدد الجزيئات لكل حويصلة قطرها 100 أو 200 أو 400 نانومتر. أ: عندما تكون البروتينات المغلفة (الإنزيمات، المجسات) في نطاق 1-10 ميكرومتر. (ب) تتم إعادة التكوين عند 1 إلى 1000 ، ومن 1 إلى 10000 ، ومن 1 إلى 10000 ، ومن 1 إلى 100000 بروتين غشائي لكل ليبيد (مول / مول). نفترض أن الجزيئات مغلفة بالتركيزات المشار إليها ومدمجة في الغشاء بنسب البروتين إلى الدهون هذه. بالنسبة لبعض الإنزيمات، رأينا أنها ترتبط بالأغشية، وهو ما قد يزيد من تركيزها الظاهري في الحويصلات. اختصار: LPR = نسبة البروتين الدهني الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
