Il campo della biologia sintetica bottom-up si concentra sulla costruzione di cellule (minime) 1,2 e bioreattori metabolici per scopi biotecnologici 3,4 o biomedici 5,6,7,8. La costruzione di cellule sintetiche fornisce una piattaforma unica che consente ai ricercatori di studiare le proteine (di membrana) in condizioni ben definite che imitano quelle degli ambienti nativi, consentendo la scoperta di proprietà emergenti e funzioni biochimiche nascoste delle proteine e delle reti di reazione9. Come passo intermedio verso una cellula sintetica funzionante in modo autonomo, vengono sviluppati moduli che catturano le caratteristiche essenziali delle cellule viventi come la conservazione dell’energia metabolica, la sintesi di proteine e lipidi e l’omeostasi. Tali moduli non solo migliorano la nostra comprensione della vita, ma hanno anche potenziali applicazioni nei campi della medicina8 e della biotecnologia10.
Le proteine transmembrana sono al centro di praticamente qualsiasi rete metabolica in quanto trasportano molecole dentro o fuori dalla cellula, segnalano e rispondono alla qualità dell’ambiente e svolgono numerosi ruoli biosintetici. Pertanto, l’ingegnerizzazione di moduli metabolici in cellule sintetiche richiede nella maggior parte dei casi la ricostituzione di proteine di membrana integrali e/o periferiche in un doppio strato di membrana composto da lipidi specifici e ad alta integrità (bassa permeabilità). La gestione di queste proteine di membrana è impegnativa e richiede conoscenze specifiche e capacità sperimentali.
Sono stati sviluppati diversi metodi per ricostituire le proteine di membrana all’interno delle vescicole fosfolipidiche, il più delle volte con lo scopo di studiare la funzione11,12, la regolazione13, le proprietà cinetiche14,15, la dipendenza dai lipidi15,16 e/o la stabilità17 di una specifica proteina. Questi metodi comportano la rapida diluizione delle proteine solubilizzate con detergente in mezzi acquosi in presenza di lipidi18, la rimozione dei detergenti mediante incubazione di proteine solubilizzate con detergente con vescicole lipidiche destabilizzate con detergente e l’assorbimento del detergente o dei detergenti su perle di polistirene19, o la rimozione dei detergenti mediante dialisi o cromatografia ad esclusione dimensionale20. I solventi organici sono stati utilizzati per formare vescicole lipidiche, ad esempio, attraverso la formazione di interfasi olio-acqua21, ma la maggior parte delle proteine integrali di membrana sono inattivate quando esposte a tali solventi.
Nel nostro laboratorio, ricostituiamo per lo più le proteine di membrana con il metodo dell’assorbimento dei detergenti per formare grandi vescicole unilamellari (LUV)19. Questo metodo consente la co-ricostituzione di più proteine di membrana e l’incapsulamento nel lume della vescicola di enzimi, metaboliti e sonde22,23. I LUV contenenti proteine di membrana possono essere convertiti in vescicole unilamellari giganti (GUV) con/senza incapsulamento di componenti idrosolubili, utilizzando l’elettroformazione24 o il rigonfiamento assistito da gel25 e condizioni specifiche per preservare l’integrità delle proteine di membrana26.
Questo articolo presenta un protocollo per la ricostituzione nei LUV di una rete metabolica fuori equilibrio che rigenera l’ATP attraverso la scomposizione della L-arginina in L-ornitina27. La formazione di ATP è accoppiata alla produzione di glicerolo-3-fosfato (G3P), un importante elemento costitutivo per la sintesi dei fosfolipidi22,28. La via metabolica è costituita da due proteine integrali di membrana, un’arginina/ornitina (ArcD) e un antiporter G3P/Pi (GlpT). Inoltre, sono necessari tre enzimi solubili (ArcA, ArcB, ArcC) per il riciclaggio dell’ATP e il GlpK viene utilizzato per convertire il glicerolo in glicerolo-3-fosfato, utilizzando l’ATP dalla scomposizione della L-arginina, vedere la Figura 1 per una panoramica schematica del percorso. Questo protocollo rappresenta un buon punto di partenza per la futura costruzione di reti di reazione ancora più complesse, per la sintesi di lipidi o proteine o per la divisione delle cellule. La composizione lipidica delle vescicole supporta l’attività di un’ampia varietà di proteine integrali di membrana ed è stata ottimizzata per il trasporto di diverse molecole dentro o fuori le vescicole 27,29,30.
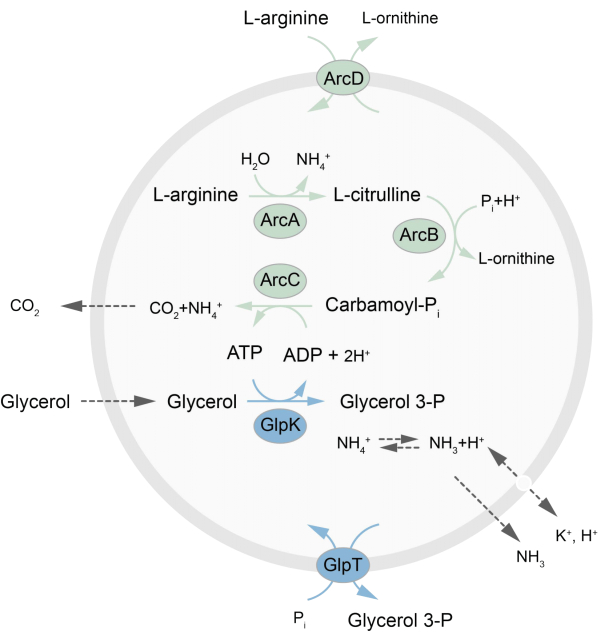
Figura 1: Panoramica della via per la produzione di ATP e la sintesi e l’escrezione del glicerolo-3-fosfato. Clicca qui per visualizzare una versione più grande di questa figura.
In breve, le proteine di membrana purificate (solubilizzate in dodecil-β-D-maltoside, DDM) vengono aggiunte alle vescicole lipidiche preformate che sono state destabilizzate con Triton X-100, che consente l’inserimento delle proteine nella membrana. Le molecole detergenti vengono successivamente (lentamente) rimosse mediante l’aggiunta di perle di polistirene attivate, con conseguente formazione di proteoliposomi ben sigillati. I componenti solubili possono quindi essere aggiunti alle vescicole e incapsulati tramite cicli di congelamento-scongelamento, che intrappolano le molecole nel processo di fusione della membrana. Le vescicole ottenute sono altamente eterogenee e molte sono multilamellari. Vengono quindi estrusi attraverso un filtro in policarbonato con una dimensione dei pori di 400, 200 o 100 nm, che produce vescicole di dimensioni più uniformi; Più piccola è la dimensione dei pori, più omogenee e unilamellari sono le vescicole ma al prezzo di un volume interno inferiore. Le proteine non incorporate e le piccole molecole vengono rimosse dalla soluzione esterna mediante cromatografia ad esclusione dimensionale. I proteoLUV possono essere convertiti in vescicole di dimensioni micrometriche mediante rigonfiamento assistito da gel, e questi proteoGUV vengono quindi raccolti e intrappolati in un chip microfluidico per la caratterizzazione e la manipolazione microscopica. La Figura 2 mostra una panoramica schematica dell’intero protocollo.
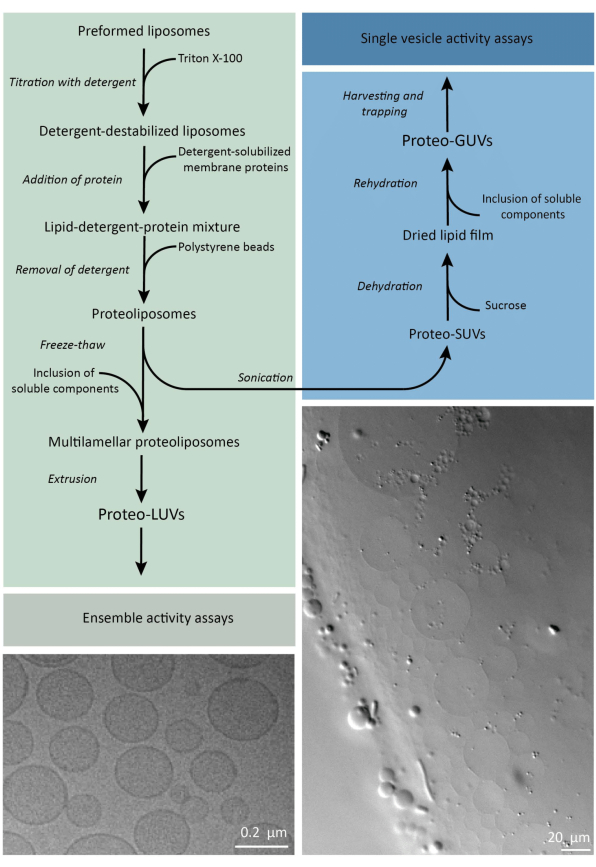
Figura 2: Panoramica del protocollo per la ricostituzione delle proteine di membrana e l’incapsulamento degli enzimi e dei componenti idrosolubili in vescicole lipidiche di dimensioni sub-micrometriche (LUV) e micrometriche (GUV). Fare clic qui per visualizzare una versione più grande di questa figura.
I protocolli di ricostituzione e incapsulamento funzionano bene e la funzionalità delle proteine viene mantenuta, ma i proteoLUV e i proteoGUV sono di dimensioni eterogenee. Gli approcci microfluidici31,32 consentono la formazione di vescicole di dimensioni micrometriche di dimensioni più omogenee, ma la ricostituzione funzionale delle proteine di membrana non è generalmente possibile perché il solvente residuo nel doppio strato inattiva le proteine. Le dimensioni dei proteoLUV variano da 100 a 400 nm e, a basse concentrazioni di enzimi, l’incapsulamento può portare a vescicole con vie metaboliche incomplete (effetti stocastici; vedi Figura 3). I LUV sono ideali per la costruzione di specifici moduli metabolici, come mostrato qui per la produzione di ATP e di elementi costitutivi come il G3P. Tali proteoLUV possono potenzialmente essere incapsulati in GUV e fungere da compartimenti simili a organelli per le vescicole dell’ospite.
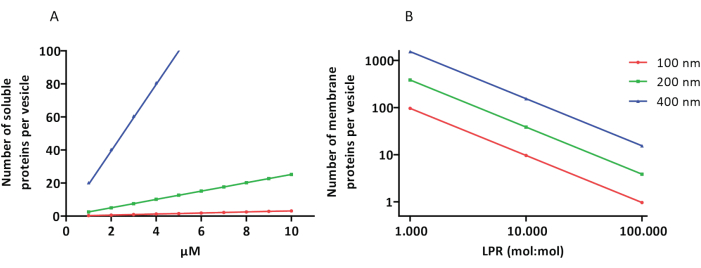
Figura 3: Numero di molecole per vescicola con un diametro di 100, 200 o 400 nm. (A) Quando le proteine incapsulate (enzimi, sonde) sono nell’intervallo 1-10 μM. (B) La ricostituzione viene eseguita a 1-1.000, da 1 a 10.000 e da 1 a 100.000 proteine di membrana per lipide (mol/mol). Partiamo dal presupposto che le molecole siano incapsulate alle concentrazioni indicate e incorporate nella membrana a questi rapporti proteina-lipide. Per alcuni enzimi, abbiamo visto che si legano alle membrane, che possono aumentare la loro concentrazione apparente nelle vescicole. Abbreviazione: LPR = Rapporto lipidico-proteico Fare clic qui per visualizzare una versione più grande di questa figura.
