Feltet nedenfra og opp syntetisk biologi fokuserer på å konstruere (minimale) celler 1,2 og metabolske bioreaktorer for bioteknologiske 3,4 eller biomedisinske formål 5,6,7,8. Konstruksjonen av syntetiske celler gir en unik plattform som lar forskere studere (membran) proteiner under veldefinerte forhold som etterligner de i innfødte miljøer, noe som muliggjør oppdagelsen av fremvoksende egenskaper og skjulte biokjemiske funksjoner til proteiner og reaksjonsnettverk9. Som et mellomtrinn mot en autonomt fungerende syntetisk celle, utvikles moduler som fanger opp essensielle trekk ved levende celler som metabolsk energibevaring, protein- og lipidsyntese og homeostase. Slike moduler forbedrer ikke bare vår forståelse av livet, men har også potensielle anvendelser innen medisin8 og bioteknologi10.
Transmembrane proteiner er kjernen i praktisk talt ethvert metabolsk nettverk når de transporterer molekyler inn eller ut av cellen, signaliserer og reagerer på kvaliteten på miljøet, og spiller en rekke biosyntetiske roller. Dermed krever konstruksjonen av metabolske moduler i syntetiske celler i de fleste tilfeller rekonstituering av integrerte og/eller perifere membranproteiner til et membrandobbeltlag sammensatt av spesifikke lipider og høy integritet (lav permeabilitet). Håndteringen av disse membranproteinene er utfordrende og krever spesifikk kunnskap og eksperimentelle ferdigheter.
Flere metoder er utviklet for å rekonstituere membranproteiner i fosfolipidvesikler, oftest med det formål å studere funksjonen11,12, regulering13, kinetiske egenskaper14,15, lipidavhengighet15,16 og/eller stabilitet17 til et spesifikt protein. Disse metodene involverer rask fortynning av vaskemiddeloppløst protein til vandige medier i nærvær av lipider18, fjerning av vaskemidler ved å inkubere vaskemiddeloppløselig protein med vaskemiddeldestabiliserte lipidvesikler og absorpsjon av vaskemiddel(e) på polystyrenperler19, eller fjerning av vaskemidler ved dialyse eller størrelseseksklusjonskromatografi20. Organiske løsningsmidler har blitt brukt til å danne lipidvesikler, for eksempel via dannelse av olje-vann-interfaser21, men flertallet av integrerte membranproteiner inaktiveres når de utsettes for slike løsningsmidler.
I laboratoriet vårt rekonstituerer vi for det meste membranproteiner ved vaskemiddelabsorpsjonsmetoden for å danne store unilamellære vesikler (LUV)19. Denne metoden tillater samrekonstituering av flere membranproteiner og innkapsling i vesikkellumen av enzymer, metabolitter og sonder22,23. De membranproteinholdige LUV-ene kan omdannes til gigantiske unilamellære vesikler (GUV) med/uten innkapsling av vannløselige komponenter, ved bruk av enten elektroformasjon24 eller gelassistert hevelse25 og spesifikke forhold for å bevare integriteten til membranproteinene26.
Denne artikkelen presenterer en protokoll for rekonstituering i LUV-er av et metabolsk nettverk utenfor likevekt som regenererer ATP gjennom nedbrytning av L-arginin til L-ornitin27. Dannelsen av ATP er koblet til produksjonen av glyserol-3-fosfat (G3P), en viktig byggestein for fosfolipidsyntese22,28. Den metabolske veien består av to integrerte membranproteiner, en arginin/ornitin (ArcD) og en G3P/Pi-antiporter (GlpT). I tillegg kreves tre løselige enzymer (ArcA, ArcB, ArcC) for resirkulering av ATP, og GlpK brukes til å omdanne glyserol til glyserol 3-fosfat, ved bruk av ATP fra nedbrytningen av L-arginin, se figur 1 for en skjematisk oversikt over banen. Denne protokollen representerer et godt utgangspunkt for fremtidig konstruksjon av enda mer komplekse reaksjonsnettverk – for syntese av lipider eller proteiner eller deling av celler. Lipidsammensetningen til vesiklene støtter aktiviteten til et bredt utvalg av integrerte membranproteiner og er optimalisert for transport av forskjellige molekyler inn i eller ut av vesiklene 27,29,30.
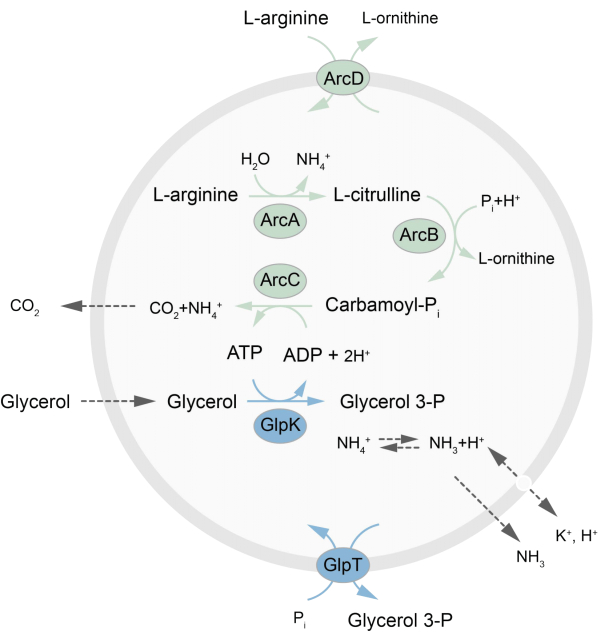
Figur 1: Oversikt over veien for ATP-produksjon og syntese og utskillelse av glyserol 3-fosfat, klikk her for å se en større versjon av denne figuren.
Kort sagt, rensede membranproteiner (oppløst i dodecyl-β-D-maltosid, DDM) tilsettes til forhåndsformede lipidvesikler som har blitt destabilisert med Triton X-100, noe som gjør det mulig å sette inn proteinene i membranen. Vaskemiddelmolekylene fjernes deretter (sakte) ved tilsetning av aktiverte polystyrenperler, noe som resulterer i dannelse av godt forseglede proteoliposomer. Løselige komponenter kan deretter tilsettes vesiklene og innkapsles via fryse-tine-sykluser, som fanger molekylene i prosessen med membranfusjon. De oppnådde vesiklene er svært heterogene og mange er multilamellære. De ekstruderes deretter gjennom et polykarbonatfilter med en porestørrelse på 400, 200 eller 100 nm, noe som gir mer jevne vesikler; Jo mindre porestørrelsen er, desto mer homogene og unilamellære er vesiklene, men til prisen av et mindre indre volum. Ikke-inkorporerte proteiner og små molekyler fjernes fra den eksterne løsningen ved størrelseseksklusjonskromatografi. ProteoGUL-ene kan omdannes til vesikler i mikrometerstørrelse ved gelassistert hevelse, og disse proteoGUVene blir deretter samlet og fanget i en mikrofluidisk brikke for mikroskopisk karakterisering og manipulasjon. Figur 2 viser en skjematisk oversikt over hele protokollen.
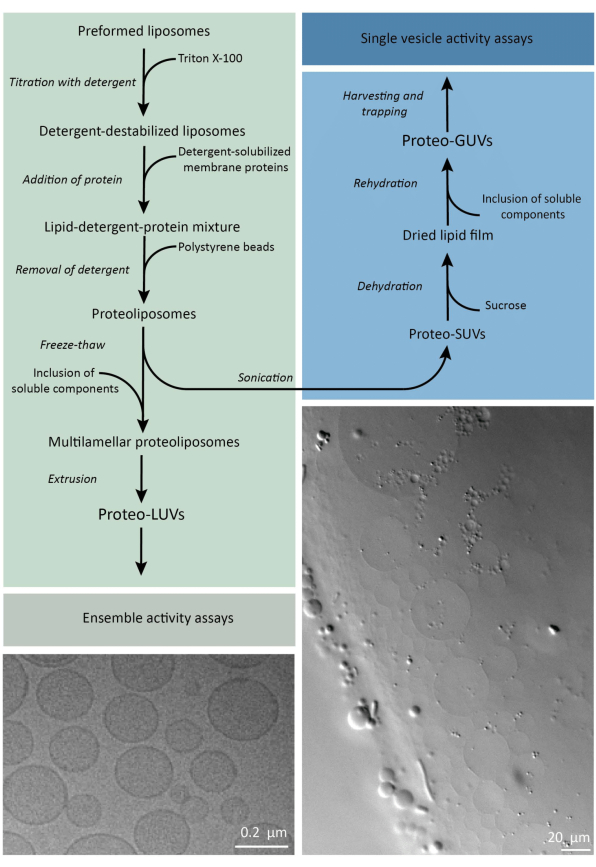
Figur 2: Oversikt over protokollen for rekonstituering av membranproteiner og innkapsling av enzymer og vannløselige komponenter i lipidvesikler av submikrometer (UV) og mikrometerstørrelse (GUV). Klikk her for å se en større versjon av denne figuren.
Rekonstituerings- og innkapslingsprotokollene fungerer bra og funksjonaliteten til proteinene beholdes, men proteoGUL-ene og proteoGUV-ene er heterogene i størrelse. Mikrofluidiske tilnærminger 31,32 tillater dannelse av vesikler i mikrometerstørrelse som er mer homogene i størrelse, men funksjonell rekonstituering av membranproteiner er generelt ikke mulig fordi gjenværende løsningsmiddel i dobbeltlaget inaktiverer proteinene. ProteoLÚVene varierer i størrelse fra 100 til 400 nm, og ved lave konsentrasjoner av enzymer kan innkapslingen føre til vesikler med ufullstendige metabolske veier (stokastiske effekter; se figur 3). LUV-er er ideelle for å konstruere spesifikke metabolske moduler, som vist her for produksjon av ATP og byggesteiner som G3P. Slike proteoLIV-er kan potensielt innkapsles i GUV-er og tjene som organelllignende rom for vertsvesiklene.
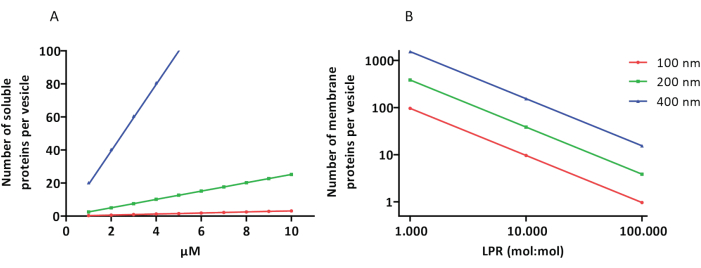
Figur 3: Antall molekyler per vesikkel med en diameter på 100, 200 eller 400 nm. (A) Når de innkapslede proteinene (enzymer, prober) er i området 1-10 μM. (B) Rekonstitueringen gjøres ved 1 til 1 000, 1 til 10 000 og 1 til 100 000 membranproteiner per lipid (mol/mol). Vi antar at molekyler er innkapslet i de angitte konsentrasjonene og inkorporert i membranen ved disse protein-til-lipid-forholdene. For noen enzymer har vi sett at de binder seg til membraner, noe som kan øke deres tilsynelatende konsentrasjon i vesiklene. Forkortelse: LPR = Lipid-Protein-Ratio Klikk her for å se en større versjon av denne figuren.
