O campo da biologia sintética de baixo para cima concentra-se na construção de células (mínimas) 1,2 e biorreatores metabólicos para fins biotecnológicos 3,4 ou biomédicos 5,6,7,8. A construção de células sintéticas fornece uma plataforma única que permite aos pesquisadores estudar proteínas (de membrana) em condições bem definidas, imitando as de ambientes nativos, permitindo a descoberta de propriedades emergentes e funções bioquímicas ocultas de proteínas e redes de reação9. Como um passo intermediário em direção a uma célula sintética que funciona de forma autônoma, são desenvolvidos módulos que capturam características essenciais das células vivas, como conservação de energia metabólica, síntese de proteínas e lipídios e homeostase. Esses módulos não apenas aprimoram nossa compreensão da vida, mas também têm aplicações potenciais nos campos da medicina8 e da biotecnologia10.
As proteínas transmembranares estão no centro de praticamente qualquer rede metabólica, pois transportam moléculas para dentro ou para fora da célula, sinalizam e respondem à qualidade do ambiente e desempenham vários papéis biossintéticos. Assim, a engenharia de módulos metabólicos em células sintéticas requer, na maioria dos casos, a reconstituição de proteínas integrais e/ou periféricas da membrana em uma bicamada de membrana composta por lipídios específicos e de alta integridade (baixa permeabilidade). O manuseio dessas proteínas de membrana é desafiador e requer conhecimentos específicos e habilidades experimentais.
Vários métodos foram desenvolvidos para reconstituir proteínas de membrana dentro de vesículas fosfolipídicas, na maioria das vezes com o objetivo de estudar a função11,12, regulação13, propriedades cinéticas14,15, dependência lipídica15,16 e/ou estabilidade17 de uma proteína específica. Esses métodos envolvem a rápida diluição da proteína solubilizada em detergente em meio aquoso na presença de lipídios18, a remoção de detergentes pela incubação de proteínas solubilizadas em detergente com vesículas lipídicas desestabilizadas por detergente e a absorção do(s) detergente(s) em esferas de poliestireno19, ou a remoção de detergentes por diálise ou cromatografia de exclusão de tamanho20. Solventes orgânicos têm sido usados para formar vesículas lipídicas, por exemplo, através da formação de interfases óleo-água21, mas a maioria das proteínas integrais da membrana são inativadas quando expostas a esses solventes.
Em nosso laboratório, reconstituímos principalmente proteínas de membrana pelo método de absorção de detergente para formar grandes vesículas unilamelares (LUVs) 19 . Esse método permite a co-reconstituição de múltiplas proteínas de membrana e a encapsulação no lúmen da vesícula de enzimas, metabólitos e sondas22,23. As LUVs contendo proteínas de membrana podem ser convertidas em vesículas unilamelares gigantes (GUVs) com/sem encapsulamento de componentes solúveis em água, usando eletroformação24 ou inchaço assistido por gel25 e condições específicas para preservar a integridade das proteínas de membrana26.
Este trabalho apresenta um protocolo para a reconstituição em LUVs de uma rede metabólica fora de equilíbrio que regenera ATP através da quebra de L-arginina em L-ornitina27. A formação de ATP está associada à produção de glicerol-3-fosfato (G3P), um importante bloco de construção para a síntese de fosfolipídios22,28. A via metabólica consiste em duas proteínas integrais da membrana, uma arginina/ornitina (ArcD) e um antiportador G3P/Pi (GlpT). Além disso, três enzimas solúveis (ArcA, ArcB, ArcC) são necessárias para a reciclagem de ATP, e GlpK é usado para converter glicerol em glicerol 3-fosfato, usando o ATP da quebra de L-arginina, consulte a Figura 1 para uma visão geral esquemática da via. Este protocolo representa um bom ponto de partida para a futura construção de redes de reação ainda mais complexas – para a síntese de lipídios ou proteínas ou a divisão de células. A composição lipídica das vesículas suporta a atividade de uma ampla variedade de proteínas integrais da membrana e foi otimizada para o transporte de diversas moléculas para dentro ou para fora das vesículas 27,29,30.
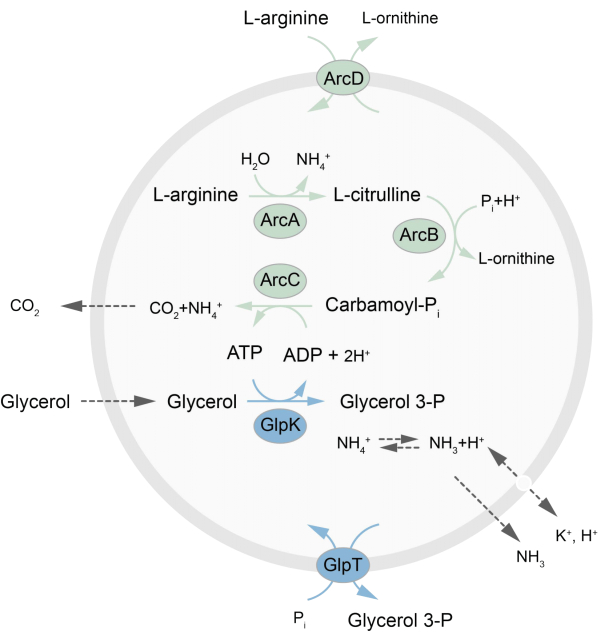
Figura 1: Visão geral da via de produção de ATP e síntese e excreção de glicerol 3-fosfato. Clique aqui para ver uma versão maior desta figura.
Em suma, proteínas de membrana purificadas (solubilizadas em dodecil-β-D-maltosídeo, DDM) são adicionadas a vesículas lipídicas pré-formadas que foram desestabilizadas com Triton X-100, o que permite a inserção das proteínas na membrana. As moléculas de detergente são subsequentemente (lentamente) removidas pela adição de esferas de poliestireno ativadas, resultando na formação de proteolipossomas bem selados. Os componentes solúveis podem então ser adicionados às vesículas e encapsulados por meio de ciclos de congelamento e descongelamento, que prendem as moléculas no processo de fusão da membrana. As vesículas obtidas são altamente heterogêneas e muitas são multilamelares. Eles são então extrudados através de um filtro de policarbonato com um tamanho de poro de 400, 200 ou 100 nm, o que produz vesículas de tamanho mais uniforme; Quanto menor o tamanho dos poros, mais homogêneas e unilamelares são as vesículas, mas ao preço de um volume interno menor. As proteínas não incorporadas e as pequenas moléculas são removidas da solução externa por cromatografia de exclusão por tamanho. Os proteoLUVs podem ser convertidos em vesículas de tamanho micrométrico por inchaço assistido por gel, e esses proteoGUVs são então coletados e presos em um chip microfluídico para caracterização e manipulação microscópica. A Figura 2 mostra uma visão geral esquemática do protocolo completo.
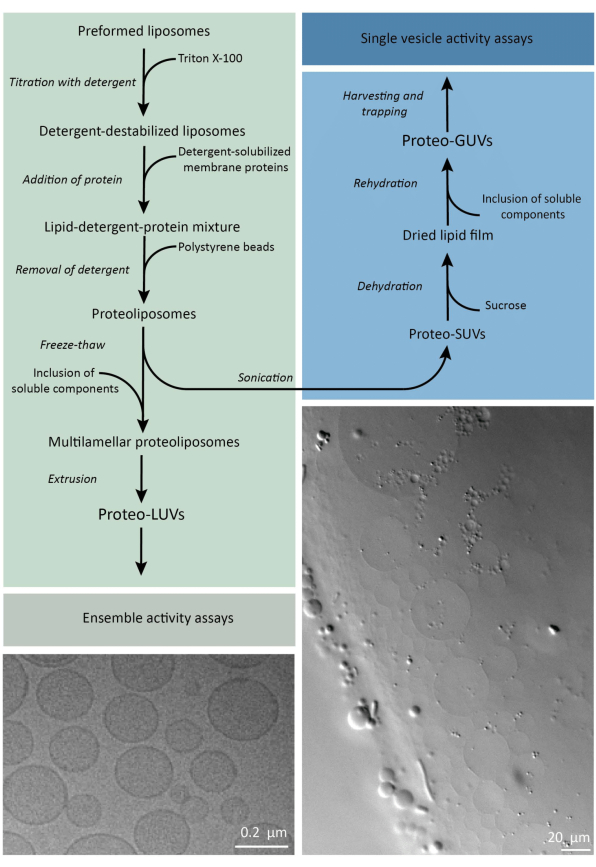
Figura 2: Visão geral do protocolo para reconstituição de proteínas de membrana e encapsulamento de enzimas e componentes solúveis em água em vesículas lipídicas de tamanho submicrométrico (LUVs) e micrométrico (GUVs). Clique aqui para ver uma versão maior desta figura.
Os protocolos de reconstituição e encapsulamento funcionam bem e a funcionalidade das proteínas é mantida, mas os proteoLUVs e proteoGUVs são heterogêneos em tamanho. As abordagens microfluídicas31,32 permitem a formação de vesículas do tamanho de micrômetros que são mais homogêneas em tamanho, mas a reconstituição funcional das proteínas da membrana geralmente não é possível porque o solvente residual na bicamada inativa as proteínas. Os proteoLUVs variam em tamanho de 100 a 400 nm e, em baixas concentrações de enzimas, o encapsulamento pode levar a vesículas com vias metabólicas incompletas (efeitos estocásticos; ver Figura 3). Os LUVs são ideais para a construção de módulos metabólicos específicos, como mostrado aqui para a produção de ATP e blocos de construção como G3P. Tais proteoLUVs podem potencialmente ser encapsulados em GUVs e servir como compartimentos semelhantes a organelas para as vesículas hospedeiras.
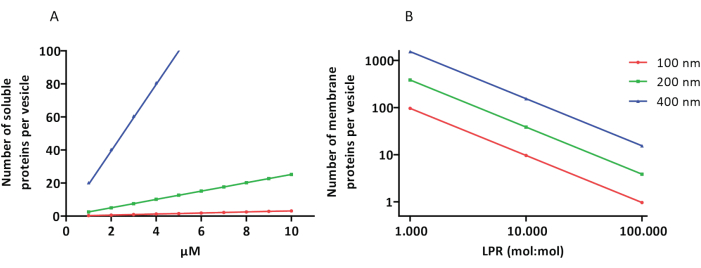
Figura 3: Número de moléculas por vesícula com diâmetro de 100, 200 ou 400 nm. (A) Quando as proteínas encapsuladas (enzimas, sondas) estão na faixa de 1-10 μM. (B) A reconstituição é feita em 1 a 1.000, 1 a 10.000 e 1 a 100.000 proteínas de membrana por lipídio (mol / mol). Assumimos que as moléculas são encapsuladas nas concentrações indicadas e incorporadas na membrana nessas proporções proteína-lipídio. Para algumas enzimas, vimos que elas se ligam às membranas, o que pode aumentar sua concentração aparente nas vesículas. Abreviatura: LPR = Proporção Lipídio-Proteína Clique aqui para ver uma versão maior desta figura.
