Staining of the lingual epithelium with antibodies to dsRed and keratin-8 (a general taste-bud marker) labeled both whole taste buds and all taste-bud innervation in Phox2b-Cre:tdTomato mice50,51 (Figure 3A). Imaging these taste buds from their pores to their bases gave the highest resolution x-y plane images (Figure 3A,B). The contour function of the pixel-based imaging program was used to outline the periphery of the taste bud in each section (Figure 3B), and then generate a surface (Figure 3C) that represented taste bud volume. Masking (or duplicating) the fluorescence associated with the taste-bud label only within the surface created a new channel that contained only this fluorescence and eliminated any papilla staining obscuring the taste bud (Figure 3D). The nerve fiber fluorescence within the taste bud was masked (Figure 3E) and used to automatically create a surface representing the volume of innervation within it (Figure 3F). A similar approach was also used to measure taste-bud volume and that of its associated innervation in circumvallate taste buds (Figure 3G). Representative measurement data revealed no correlations between taste-bud volumes and innervation volumes in either the fungiform (p = 0.115) or the circumvallate (p = 0.090) measurement regions (Figure 3H).
The administration of a low dose of tamoxifen in TrkBCreER:tdTomato mice causes gene recombination and the labeling of a small number of neurons so that taste buds are innervated by zero to a few labeled terminal arbors (the neuronal portion within the taste bud). The lingual epithelium was stained using an anti-dsRed antibody for the terminal arbors and anti-Car4 (sour) and anti-PLCβ2 (sweet, bitter, and umami) antibodies for the taste-transducing cells (Figure 4A). A vector-based image analysis program was used to trace the labeled terminal arbors (Figure 4B). The orthogonal heights of the arbors associated with the blue and green tracings were 33.4 µm (Figure 4C) and 32.4 µm (Figure 4D), respectively. The 3D Convex Hull measurements (i.e., the extent of the terminal arbor within the taste bud) for the blue terminal arbor was 644.0 µm3 and 3647.0 µm3 for the green arbor. The dendrogram for the green tracing is shown in Figure 4E with branch lengths measured in microns. The green arbor had seven branch ends and a total length of 183.4 µm. Quantification of the absolute numbers of PLCβ2+ and Car4+ cells revealed that this taste bud had 17 PLCβ2+ cells and two Car4+ cells. Using cell pixel-based imaging software to determine the closest proximity between nerve fibers and taste-transducing cells revealed that out of a total of 19 taste-transducing cells in the taste bud, the blue terminal arbor (shown in red in Figure 4F,G) was within 200 nm (the resolution of the light microscope) of the light blue Car4+ cell (white areas indicated by arrows in Figure 4G). The terminal arbor associated with the green tracing is shown in magenta (Figure 4F and Figure 4H) and is within 200 nm of both the light and dark blue Car4+ cells (white areas in Figure 4H). As the next closest cell to these arbors was more than 200 nm away, there was an unlabeled voxel separating the two structures.
Dividing progenitor cells were labeled using injections of EdU on Days 0, 1, and 3, and tissues were collected on Day 4. Whole-mount keratin-8 and EdU staining of fungiform taste buds revealed that EdU-labeled cells were present both within and outside of the taste buds (Figure 5A–C). Individual EdU+/keratin-8+ cells (teal and yellow) and EdU+/keratin-8- nuclei (purple and magenta) were segmented (Figure 5B,C). The dark blue cell shown was keratin-8+ and had an elongated shape consistent with mature taste-transducing cells. These surfaces are shown with the taste bud oriented from pore-to-base (Figure 5B) and along the long axis of the taste bud (Figure 5C). Each structure could be viewed in individual optical slices by masking the fluorescence within each structure (Figure 5D–F). The magenta and purple nuclei are outside of the keratin-8+ border of the taste bud indicated by the white-dotted outline (Figure 5D,E). The yellow, teal, and blue cells were within the taste bud (Figure 5D–F). Individual taste-transducing cells could be reconstructed using pixel-based imaging software of either Car4 labeling (Figure 6A–C) or PLCβ2 labeling (Figure 6D–F). A pixel-based imaging software was be used to measure the closest proximity between cells revealed that a Car4+ cell (same cell as shown in Figure 6B) was within 200 nm of a single PLCβ2+ cell (Figure 6G, green). The area where the cells were within 200 nm of each other is shown in white (Figure 6G) and indicated by white arrows. The next closest cell was more than 200 nm away and is shown in yellow in Figure 6H,I in two different orientations. Figure 7 demonstrates the isolation and analysis of the innervation terminating within the papilla (but outside the taste bud) and includes its distribution around the taste bud and its distance from the epithelium.
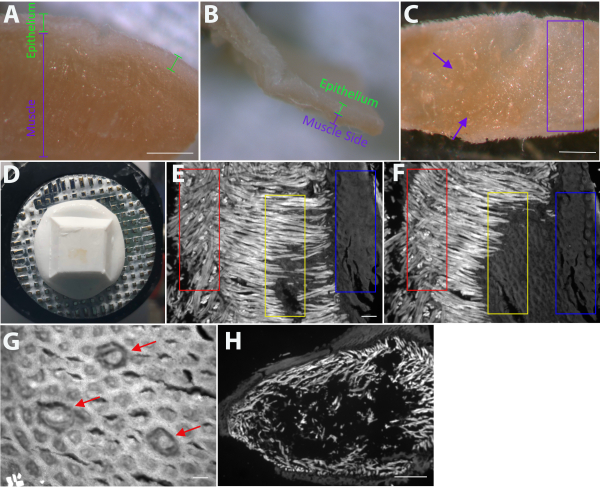
Figure 1: Preparation of lingual epithelium for fungiform taste-bud staining. (A) View of the cut tongue with epithelium and muscle labeled prior to any dissection. (B) Once enough muscle has been removed, there is only a small amount of remaining muscle on the underside of the epithelium. In addition to evaluating the progress of the dissection by viewing the cut side of the epithelium, (C) laying the epithelium flat on a glass slide under the dissecting scope reveals that some portions of the tissue are evenly translucent (purple rectangle); enough muscle has been removed from this area. In contrast, the purple arrows indicate regions on the left where there is more muscle that needs to be removed. Once the entire underside of the epithelium is similar to the area in the purple rectangle, proceed to the next step. (D) After portions of the epithelium have been frozen with the muscle side down, additional muscle and lamina propria are removed as thin sections using the cryostat. When sectioning is complete, the remaining epithelium is thin and translucent. (E–F) Serial sections (20 µm) were collected on a glass slide, and each section was viewed under a fluorescent microscope before cutting the next section. Well below the epithelium, muscle fibers are oriented in multiple directions so that muscle fibers are present both in cross section and along the muscle fiber (E, red rectangle). The serial sections in E–F demonstrate the transition from muscle fibers oriented in multiple directions (E, red rectangle) to muscle fibers being oriented mostly in one direction (F, red rectangle), which is indicative of the muscle-lamina propria border. Another region of the same piece of tissue (yellow rectangles) demonstrates that when the muscle fibers are oriented in one direction, the next section will likely yield connective tissue because all muscle has been removed from that region. The blue rectangles both represent the underside of the epithelium. If taste buds are present on the section (G, red arrows), too much tissue has been removed. Ideally, sectioning is complete when the underside of the epithelium (but no taste buds) is visible in the removed sections (F, yellow rectangle). Although areas with muscle fibers oriented in the same direction (E, yellow rectangle and F, red rectangle) are also suitable for sectioning, areas where the muscle fibers are oriented in multiple directions (E, red rectangle) should be avoided. (G) Once sections include the underside of the epithelium/lamina propria, it is only possible to cut a few additional sections before too much of the epithelium has been removed and sections include taste buds. (H) The most common mistake is revealed by cryostat sections where epithelium is seen at the edge of the tissue, muscle is seen inside of the epithelium, and OTC/sparse muscle is present in the middle. This is most often due to not laying the tissue flat on the bottom of the tissue mold before freezing or insufficient flattening with blunt-ended forceps. Scale bars in A–C = 1 mm; scale bars in E, F, H = 100 µm; scale bar in G = 50 µm. Please click here to view a larger version of this figure.
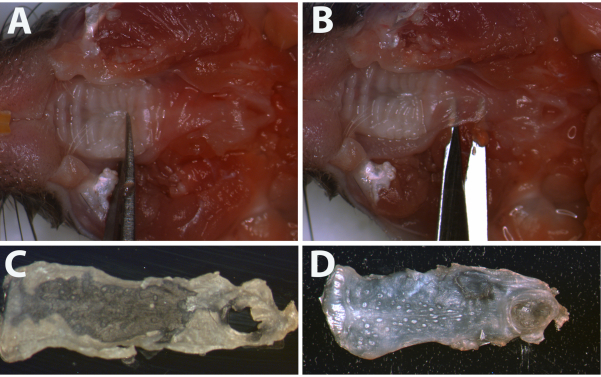
Figure 2: Dissection of palate for staining. (A) The palate was dissected first using thin blade scissors to cut the hard palate, (B) then using the same scissors to separate the soft palate from the underlying connective tissue. After removing the tissue from the oral cavity, any remaining tissue was removed with the scissors. At this point, all that may remain are glands on the back of the soft palate. A razorblade was used to gently scrape away these glands. The (C) back and (D) epithelial surface of the completed dissection of the palate are shown. Please click here to view a larger version of this figure.
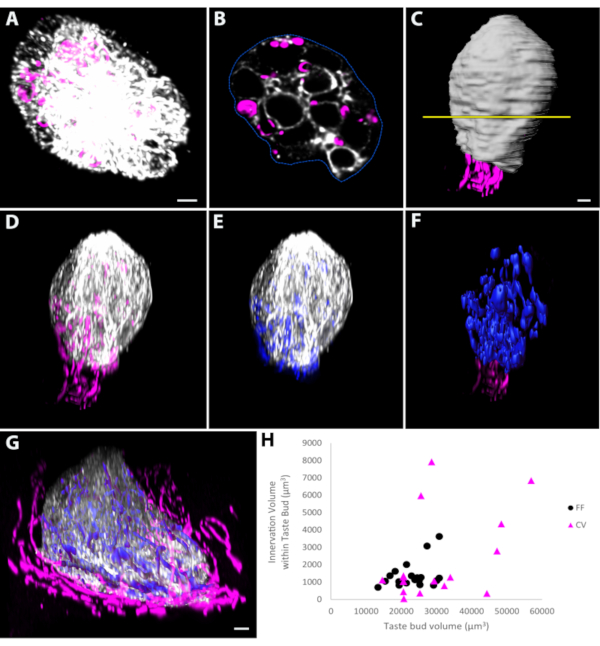
Figure 3: Measuring volume in whole-mount taste buds. (A) Whole-mount taste buds were imaged from the taste pore to the base of the taste bud so that the plane of highest resolution is the x-y plane. Each optical slice was viewed in pixel-based image analysis software, and the contour function was used to manually outline the periphery of the taste bud stained with keratin-8. (B) An example of one optical slice is provided. (C) The position of this representative section along the long axis of the taste bud is shown by the yellow line. After each optical section was outlined, a surface was created that represents the volume of the taste bud (white). Masking or duplicating the fluorescent channel corresponding to the taste bud (keratin-8 in D) or the tdTomato-labeled innervation (pseudo-colored blue in E) within the volume representing the taste bud. The fluorescence within the taste bud in (E) was used to generate a surface representing the volume of innervation within the taste bud (F, blue). (G) A similar approach was applied to whole-mount circumvallate taste buds imaged in the same orientation as the fungiform taste bud in A. (H) Measuring the volume of fungiform and circumvallate taste buds and their respective volume of innervation revealed that there is no correlation between the taste bud and innervation volume for taste buds sampled for either region. Scale bars in A–D, F = 4 µm; scale bar in G = 5 µm. This figure has been modified from Ohman-Gault et al.50. Abbreviations: FF = fungiform; CV = circumvallate. Please click here to view a larger version of this figure.
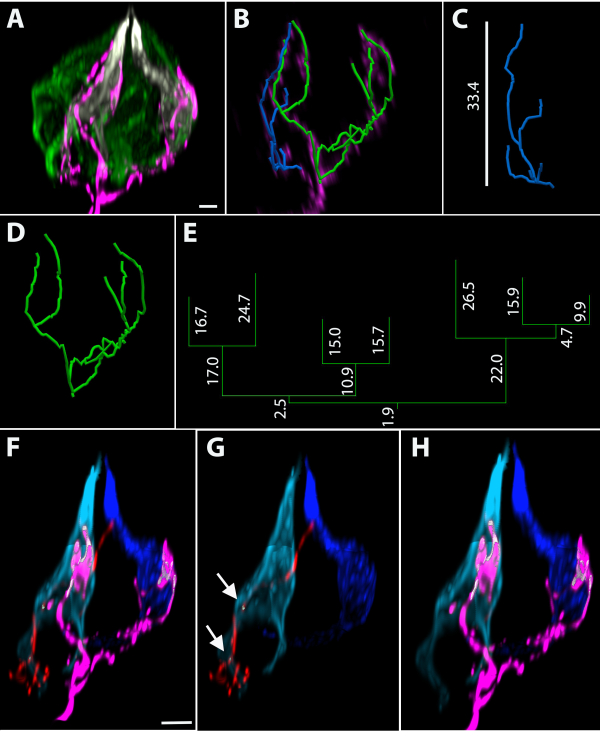
Figure 4: Representative terminal arbors in fungiform taste buds using sparse cell genetic labeling. (A) Whole-mount taste bud stained with taste-transducing-cell markers Car4 (white) and PLCβ2 (green). (B) This taste bud has two labeled terminal arbors, which are shown with the taste bud removed after reconstructing the fibers. (C) The blue arbor has 6 branch ends and an orthogonal height in the taste bud of 33.4 µm and (D) the green arbor has 7 branch ends. (E) The dendrogram corresponding to the green arbor is provided with each segment length in micrometers. (F-H) The distance between structures was measured. (F–G) The blue tracing in C was segmented and is shown in red. (G) The areas where this terminal arbor is within 200 nm of the light blue Car4+ cell are indicated by white arrows. (F, H) The terminal arbor represented by the green reconstruction is shown in magenta. (H) The magenta arbor (associated with the green tracing in 4B, D) is within 200 nm of both the dark and light blue Car4+ cells. Scale bar in A, B = 4 µm; scale bars in F–H = 5 µm. Please click here to view a larger version of this figure.
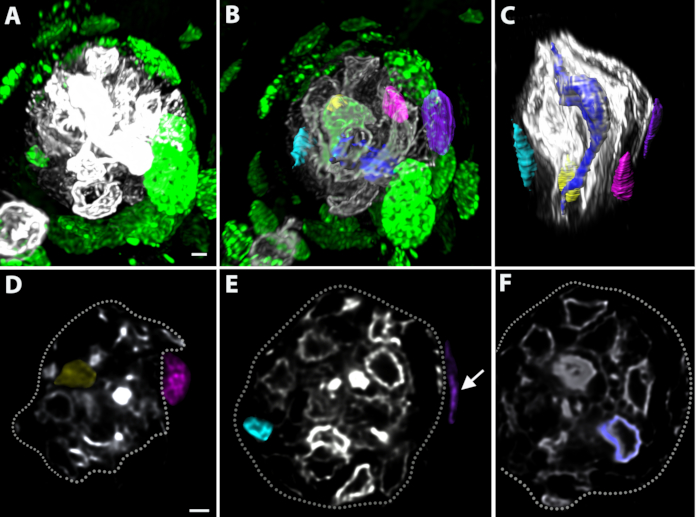
Figure 5: Whole-mounts can be used to track incorporation of new taste bud cells. Mice were injected with EdU to label dividing progenitors on Days 0, 1, and 3 and sacrificed on Day 4. (A, B) Cells labeled with EdU (green) can be identified both around and within the taste bud, which is labeled with keratin-8 (A, white, B, gray). (B, C) Individual EdU-labeled, keratin-8+ cells inside the taste bud and keratin-8-, EdU-labeled nuclei are segmented outside the taste bud. (D–F) The fluorescence within each structure segmented in A–B was masked and can be seen in cross-section. The perimeter of the taste bud is outlined with a white dotted line (D–F). (D) The yellow cell is within the taste bud and is both EdU-labeled and keratin-8+. The magenta nucleus is outside the taste bud and is keratin-8-. (E) The teal cell is inside the taste bud and both EdU-labeled and keratin-8+. The purple EdU-labeled nucleus is keratin-8- and outside of the taste bud (white arrow). (F) The blue cell is keratin-8+ and elongated, consistent with mature taste-transducing cells. Scale bars in A–C =3 µm; scale bars in D = 2 µm; scale bars in E, F = 4 µm. Abbreviation: EdU = 5-ethynyl-2′-deoxyuridine. Please click here to view a larger version of this figure.
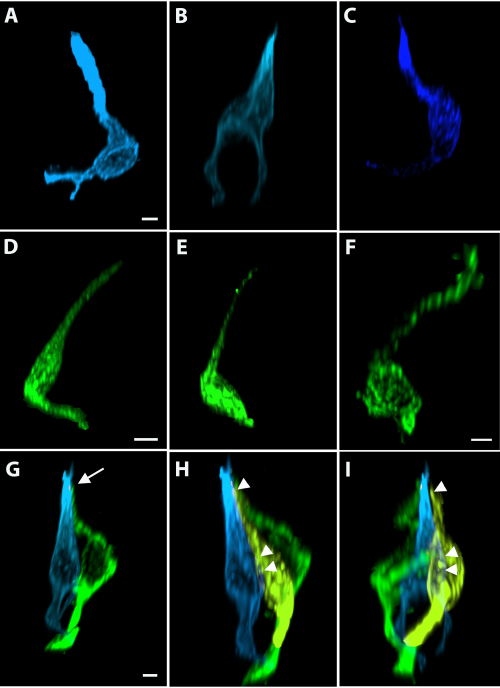
Figure 6: Shapes of whole taste bud cells can be analyzed along with their relationships with other taste bud cells. (A–F). Segmenting individual taste bud cells to create surfaces isolates individual taste bud cells, facilitating clear visualization. Individual (A–C) Car4+ and (D–F) PLCβ2+ cells show the variation in individual cell shapes. (G) The closest PLCβ2+ cell to the Car4+ cell in B was determined to be within 200 nm (at a single small 0.5 µm2 location indicated by arrow). The next closest cell was greater than 300 nm away and distinguishable as a separate structure from the segmented Car4+ cell. (H, I) The next closest cell was segmented; and the masked fluorescence is shown in yellow. The three closest points for the next closest cell (yellow)are indicated by arrowheads in H, I. Scale bars in A–C = 3 µm; scale bars in D, E = 4 µm; scale bar in F = 2 µm; scale bars in G–I = 3 µm. Please click here to view a larger version of this figure.
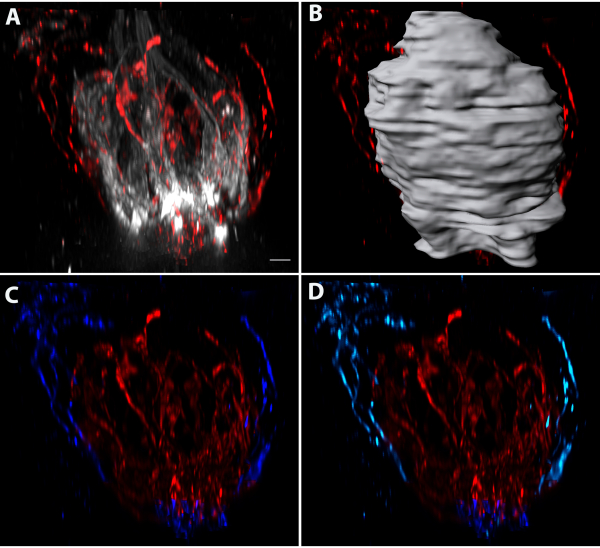
Figure 7: Quantifying innervation to the papilla. (A) Some labels for taste neurons also label innervation to the papilla.(B)The innervation within the taste bud is separated from the innervation outside the taste bud by segmenting the taste bud (as described for Figure 3), (C) masking the innervation inside the taste bud (red), and then masking the innervation outside of the taste bud only (dark blue). The volume of innervation to the taste bud (red) was 1649.6 μm3. The innervation outside the taste bud will include taste fibers underneath the papilla that should not be included in the quantification of the innervation to the papilla. (D) The fluorescence of the innervation to the papilla was masked (light blue). The volume of innervation to the papilla was 121.8 μm3. Scale bars in A-D = 4 μm. Please click here to view a larger version of this figure.
