Nous décrivons une approche d’imagerie de fluorescence et de luminosité de réflexion interne totale (TIRF-N-B) pour déterminer l’état oligomeric moyen des molécules de récepteur à la membrane de plasma des cellules vivantes, visant à relier l’assemblage de récepteur dynamique à la fonction biologique des protéines (Figure 1).
Sur la liaison extracellulaire de ligand, les récepteurs initient la transduction intracellulaire de signal selon leur conformation, oligomerization, co-récepteurs potentiels et composition de membrane. Malgré l’importance et l’omniprésence de l’oligomerisation des récepteurs, reconnu comme un événement clé dans la signalisation cellulaire1,2,3,4,5,6, 7, peu de méthodes peuvent détecter les événements de clustering et mesurer le degré de clustering expérimentalement8,9. Le volume confocal (x,y 300 nm, z ‘900 nm) est insuffisamment résolu pour prouver l’interaction moléculaire et la stoichiométrie, même après optimisation par des algorithmes de restauration d’image10. La composition sous-unité des oligomères protéiques ne peut pas être résolue sur une base purement spatiale, même par des méthodes de super-résolution à x, y résolution de 20-70 nm tels que PALM11, STORM12, et STED13. En outre, leur résolution temporelle (de l’ordre de minutes par image) ne peut pas suivre la cinétique dans la gamme de secondes. Le blanchiment par étapes d’une seule molécule ne résout la stoichiométrie des oligomères protéiques que s’ils sont immobiles14.
L’une des méthodes les plus polyvalentes pour mesurer la densité et l’oligomerisation des protéines étiquetées fluorescentes dans des images uniques est l’analyse de distribution de l’intensité spatiale (SpIDA), qui repose sur l’échantillonnage spatial. Il s’applique à la fois aux cellules fixes et vivantes chimiquement, et permet l’analyse de plusieurs régions d’intérêt de la cellule simultanément en utilisant la microscopie de fluorescence standard15. Alternativement, les méthodes de moment, telles que la spectroscopie de fluorescence-corrélation (FCS)16,l’histogramme de comptage de photon (PCH)17,et le nombre et la luminosité (N et B)18,19,sont appropriés pour l’oligomère quantitatif Mesures. Ces méthodes analysent les fluctuations d’intensité de fluorescence qui peuvent être observées à temps lorsque les fluorophores se diffusent dans et hors d’un volume d’éclairage. Les amplitudes des fluctuations d’intensité peuvent être décrites de façon unique par la luminosité moléculaire du fluorophore (mD) et le nombre moyen de fluorophores (n) dans le volume d’éclairage17 (figure 2). Typiquement, le coefficient de diffusion des fluorophores et le nombre moyen de molécules (inversement liés à la valeur G(0) dans le volume d’éclairage peuvent être obtenus par FCS20. Cependant, puisque le temps de diffusion ne s’écaille qu’avec la racine cubique de la masse, le FCS n’est pas suffisamment sensible pour détecter les changements de masse moléculaire21. Dans la pratique, FCS de couleur unique ne peut pas détecter la dimerisation des récepteurs membranaires. PCH résout avec précision les mélanges de différents oligomères. À l’aide de plus de deux moments de la distribution de l’amplitude, il détecte des molécules d’une luminosité différente qui occupent le même volume d’éclairage. Balayage FCS22 et les développements, tels que l’intéressante paire-corrélation de la luminosité moléculaire (pCOMB) approche23, introduit pour étendre la gamme d’applicabilité des méthodes de corrélation de fluorescence dans les systèmes biologiques24 , restent des méthodes à point unique dépourvues de la capacité de mesures rapides dans une grande zone d’une cellule, nécessitant de nombreuses observations consécutives à chaque pixel et acquisition de données dans l’ordre de secondes.
N-B est une version simplifiée de PCH qui ne tient compte que des premiers et deuxièmes moments de l’amplitude de la distribution de fluorescence, à savoir l’intensité moyenne, lt;I-gt;, et la variance,‘2 (Figure 2)18,19 et, pour cette raison, il ne peut pas déterminer la fraction molaire des oligomères inconnus dans un mélange, mais seulement estime l’état moyen d’oligomerisation du mélange. Néanmoins, n’a l’avantage de travailler avec des séries temporelles d’images de cellules vivantes relativement plus petites que PCH sur une base pixel par pixel, simplement en surveillant les fluctuations à l’heure de l’intensité de la fluorescence. Étant donné que le temps par pixel réduit le temps par pixel à quelques microsecondes, il peut suivre la cinétique d’oligomerisation rapide sur de grandes zones cellulaires, permettant l’acquisition d’images sur une échelle de temps de secondes dans la microscopie de balayage de raster (par exemple, confocal, 2-photon) et millisecondes microscopie à la caméra (p. ex. TIRFM).
Plusieurs rapports ont démontré la capacité de N-B à quantifier le nombre de sous-unités dans les grappes de protéines par imagerie des régions cellulaires étendues. Des amas de Paxillin-EGFP ont été détectés aux emplacements d’adhérence dans les cellules de CHO-K125,et l’agrégation intracellulaire du peptide pathogène de Httex1p a été décrite dans les cellules coS-726. Le n et B a été appliqué pour suivre l’oligomerisation ligand-conduite du récepteur d’ErbB27,et l’effet du ligand FGF21 sur Klothob (KLB) et FGFR1c dans les cellules de HeLa28. La combinaison de l’imagerie TIRF et de l’analyse N-B a été utilisée pour montrer que le dynamin-2 est principalement tétracumé dans toute la membrane cellulaire29. Nous avons appliqué n’et B à la fois raster balayage et images TIRF pour prouver ligand-conduit dimerization des récepteurs de membrane cellulaire uPAR et FGFR130,31.
Les méthodes de corrélation de fluorescence, telles que le N-B, le FCS et le PCH, sont basées sur l’idée que dans un volume ouvert le nombre d’occupation des particules suit une distribution de Poisson. Parce que seuls les photons que les fluorophores émettent peuvent être détectés, la valeur moyenne  d’une intensité de fluorescence mesurée par rapport au temps dans un pixel de l’image, est le produit du nombre moyen de fluorophores dans le volume d’éclairage, n, et leur luminosité moléculaire,17euros :
d’une intensité de fluorescence mesurée par rapport au temps dans un pixel de l’image, est le produit du nombre moyen de fluorophores dans le volume d’éclairage, n, et leur luminosité moléculaire,17euros :

où le nombre de photons émis par unité de temps (conventionnellement par seconde) par molécule lorsque la molécule est au centre du volume d’éclairage.
La luminosité est une propriété de chaque fluorophore dans une acquisition donnée mise en place, tandis que l’intensité est la somme de toutes les contributions de tous les fluorophores. Dans les concours biologiques, la luminosité augmentera avec l’augmentation du nombre de fluorophores qui fluctuent ensemble, donnant des informations sur l’état d’oligomerisation de la protéine fluorescente. Les amplitudes de fluctuation à un pixel donné sont mesurées à partir de la variance du signal de fluorescence,no 2:

Lorsque la moyenne du carré  d’intensité, et le
d’intensité, et le  carré de la moyenne de l’intensité, , sont calculer à partir des valeurs d’intensité individuelle dans chaque pixel de chaque image:
carré de la moyenne de l’intensité, , sont calculer à partir des valeurs d’intensité individuelle dans chaque pixel de chaque image:
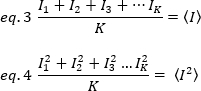
où K est le nombre total d’images dans la série chronologique. Expérimentalement, il est nécessaire de calculer pour l’ensemble de la série d’images la variance qui décrit la diffusion des valeurs d’intensité individuelles à chaque pixel d’une seule image autour de la valeur moyenne de l’intensité. La variance comprend toutes les fluctuations d’origines différentes. Dans une première approximation, la variance due aux particules de diffusion dans le volume d’éclairage,20, peut être séparée de la variance en raison du bruit de tir du détecteur,2d. Les deux écarts sont indépendants; ainsi, l’écart total est donné par leur somme :

La variance, due aux fluctuations moléculaires dans et hors du volume de détection, dépend linéairement de la luminosité et de l’intensité moléculaires :

Réorganisation eq. 6 selon eq. 1:

Selon le concept typique de la spectroscopie de corrélation de fluorescence, l’équation 7 indique que la variance due au nombre de fluctuations dépend du carré de la luminosité des particules.
Ensuite, la variance due aux fluctuations du détecteur est une fonction linéaire de l’intensité détectée, en supposant que le détecteur est exploité en dessous de sa limite de saturation19:

Dans le cas des détecteurs de comptage de photons de1 et de 0,la variance du détecteur est donc égale à l’intensité moyenne :

Pour appliquer ces concepts à des mesures réelles dans des cellules vivantes, Gratton et ses collègues18 définissent la luminosité apparente, B, pour chaque pixel comme le rapport de la variance par rapport à l’intensité moyenne :

B est le paramètre qui est mesuré expérimentalement. Dans ce travail, les images de séries chronologiques des récepteurs FGFR1 à la membrane plasmatique des cellules HeLa sont capturées par microscopie TIRF et la luminosité apparente moyenne, B, est déterminée par l’analyse N et B. Puis, après l’ajout de FGF2, des séries chronologiques consécutives sont capturées pour suivre les changements dans l’auto-assemblage des molécules de récepteur dans la surface de membrane après stimulation du récepteur avec le ligand canonique.
Cependant, puisque le détecteur du microscope TIRF est une caméra EMCCD, l’expression de la luminosité apparente doit être modifiée comme19:

lorsque le décalage est le décalage d’intensité de l’électronique de détection qui est une caractéristique des paramètres du détecteur. La variance et l’intensité moyenne d’un détecteur analogique sont données respectivement par :
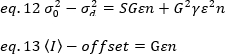
où G est le gain analogique dans les niveaux numériques (DL/photons), S, les niveaux numériques par photon19, est donné par la pente d’une intensité par rapport à l’intrigue de variance pour une source lumineuse avec une intensité constante (pas de fluctuations temporelles). Le facteur de l’aétée est lié à la forme du volume de détection des pixels. Selon Hassler et coll.32, le facteur d’aété est égal à 0,3 pour l’imagerie TIRF travaillant au gain maximum de la caméra de détection19. Les paramètres de décalage, S et G sont des caractéristiques de la caméra et du microscope. La luminosité apparente, B, est obtenue en réorganisant eq. 11 selon eq. 12 et 13:

Expérimentalement, est une fonction complexe de l’intensité laser et de l’efficacité de détection du système. Néanmoins, étant donné que le B/S dépend linéairement de l’activité, il est seulement important de déterminer la valeur relative de l’application pour un mode de détection donné :

où est proportionnelle à la proportion de ‘ Pourtant, un étalonnage est effectué à l’aide d’une référence interne.
