Мы описываем подход к визуализации Total Internal Reflection Fluorescence-Number и Brightness (TIRF-N’amp;B) для определения среднего олигомерного состояния молекул рецепторов в плазменной мембране живых клеток, направленных на увязку рецепторной сборки динамика к биологической функции белков(Рисунок 1).
При внеклеточном связывании лиганд рецепторы инициируют трансдукцию внутриклеточного сигнала в зависимости от их конформации, олигомеризации, потенциальных корецепторов и мембранного состава. Несмотря на важность и повсеместность олигомеризации рецепторов, признанных ключевым событием в клеточной сигнализации1,2,3,4,5,6, 7, несколько методов могут обнаружить события кластеризации и измерить степень кластеризации экспериментально8,9. Конфокальный объем (x,y’300 nm, z z 900 nm) недостаточно решен для доказательства молекулярного взаимодействия и стоихиометрии, даже после оптимизации алгоритмами восстановления изображения10. Подедины состав белковых олигомеров не может быть решен на чисто пространственной основе даже с помощью супер-разрешения методов на х, разрешение 20-70 нм, таких как PALM11, STORM12, и STED13. Более того, их временное разрешение (в порядке минут на изображение) не может следовать за кинетиками в диапазоне секунд. Одномолекулярная ступенчатое отбеливание разрешает стойихиометрию белковых олигомеров только в том случае, если они неподвижны14.
Одним из наиболее универсальных методов измерения плотности и олигомеризации флуоресцентно отмеченных белков в пределах одного изображения является анализ распределения пространственной интенсивности (SpIDA), который опирается на пространственную выборку. Он применим как к химически фиксированным, так и к живым клеткам, и позволяет анализировать несколько областей, представляющих интерес для клетки, одновременно с помощью стандартной флуоресценции15. Кроме того, методы момента, такие как флуоресценция-корреляция спектроскопии (FCS)16, фотон подсчета гистограммы (PCH)17, и количество и яркость (N и B)18,19, подходят для количественного олигомера Измерения. Эти методы анализируют колебания интенсивности флуоресценции, которые могут наблюдаться во времени, когда фторфоры рассеивается в и из объема освещения. Амплитуда колебаний интенсивности можно однозначно описать молекулярной яркостью флюорофора (к) и средним количеством флюорофоров (n) в пределах объема освещения17 (Рисунок 2). Как правило, коэффициент диффузии фторфоров и среднее количество молекул (обратно связанных с значением G(0) в пределах объема освещения могут быть получены FCS20. Однако, так как время диффузии только масштабируется с кубической корнем массы, FCS не является достаточно чувствительным для обнаружения изменений в молекулярной массе21. На практике, один цвет FCS не может обнаружить димеризацию мембранных рецепторов. PCH разрешает смеси различных олигомеров точно. Используя более двух мгновений распределения амплитуды, он обнаруживает молекулы разной яркости, которые занимают один и тот же объем освещения. Сканирование FCS22 и разработки, такие как интересная парно-корреляция молекулярной яркости (pCOMB) подход23, введены для расширения диапазона применимости методов корреляции флуоресценции в биологических системах24 , остаются одноточечными методами, не имеющими возможности быстрых измерений на большой площади ячейки, требующих много последовательных наблюдений на каждом пикселе и получения данных в порядке секунд.
Н З/Б является упрощенной версией PCH, которая учитывает только первый и второй моменты амплитуды распределения флуоресценции, а именно, средняя интенсивность, lt; I’gt;, и дисперсия,No 2 (Рисунок 2)18,19 и, из-за этого, он не может определить моляровую фракцию неизвестных олигомеров в смеси, а лишь оценивает среднее состояние олигомеризации смеси. Тем не менее, Н И Б имеет то преимущество, что работает с относительно меньшими временные ряды изображений живых клеток, чем PCH на пиксель за пикселем основе, просто путем мониторинга колебаний во времени флуоресценции интенсивности. Поскольку Н И Б сокращает время за пикселем до нескольких микросекунд, он может следить за быстрой кинетикой олигомеризации на больших клеточных площадях, позволяя присугнять изображение в временной шкале секунд в сканирующей микроскопии (например, confocal, 2-фотон) и миллисекундах микроскопии на основе камеры (например, TIRFM).
В нескольких докладах была продемонстрирована способность Н И Б количественно очислять количество субъединиц в белковых кластерах с помощью визуализации расширенных клеточных регионов. Кластеры Paxillin-EGFP были обнаружены на участках адгезии в клетках CHO-K125,а внутриклеточная агрегация патогенного пептида Httex1p была описана в клетках COS-726. Н З/Б был применен для следующих лиганд-управляемых олигомеризации рецептора ErbB27, и влияние лиганд FGF21 на Klothob (KLB) и FGFR1c в клетках HeLa28. Сочетание tIRF изображений и N и B анализ был использован, чтобы показать, что динамин-2 в первую очередь тетрамерические всей клеточной мембраны29. Мы применили N и B как raster сканирования и TIRF изображения доказать лиганд-управляемый димеризации uPAR и FGFR1 клеточных мембранных рецепторов30,31.
Методы корреляции флуоресценции, такие как Н И Б, FCS и PCH, основаны на том, что в открытом томе число частиц следует за распределением Пуассона. Потому что только фотоны, что фторофоры излучают могут быть обнаружены, среднее значение для измеренной интенсивности флуоресценции по сравнению со временем в пикселе изображения,  является продуктом среднего количества флюорофоров в объеме освещения,, и их молекулярная яркость, No17:
является продуктом среднего количества флюорофоров в объеме освещения,, и их молекулярная яркость, No17:

где выражено как количество фотонов, испускаемых на единицу времени (обычно в секунду) на молекулу, когда молекула находится в центре объема освещения.
Яркость является свойством каждого фторофора в данном приобретении создана, в то время как интенсивность сумма всех взносов от всех фторофоров. В биологических конкурсах, яркость будет увеличиваться с увеличением числа флюорофоров, которые колеблются вместе, давая информацию о состоянии олигомеризации флуоресцентно помечены белка. Амплитуда колебаний на данном пикселе измеряется из дисперсии сигнала флуоресценции,No 2:

Где среднее значение квадрата  интенсивности, и квадрат среднего
интенсивности, и квадрат среднего  интенсивности, , вычисляются из индивидуальных значений интенсивности в каждом пикселе каждого кадра:
интенсивности, , вычисляются из индивидуальных значений интенсивности в каждом пикселе каждого кадра:
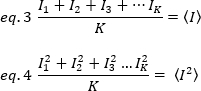
где K — это количество общих кадров в временных рядах. Экспериментально необходимо вычислить для всей серии изображений дисперсию, описывающая рассеяние значений индивидуальной интенсивности на каждом пикселе одного изображения вокруг значения средней интенсивности. Разница включает в себя все колебания разного происхождения. В первом приближении, дисперсия из-за рассеяния частиц в объеме освещения,20, могут быть отделены от дисперсии из-за шума выстрела детектора,2d. Эти два отклонения являются независимыми; таким образом, общая дисперсия дается их сумма:

Разница, из-за молекулярных колебаний в и из объема обнаружения, линейно зависит от молекулярной яркости и интенсивности:

Перестановка eq. 6 в соответствии с eq. 1:

Согласно типичной концепции флуоресценции корреляции спектроскопии, уравнение 7 гласит, что дисперсия из-за количества колебаний зависит от квадрата яркости частицы.
Затем, дисперсия из-за колебаний детектора является линейной функцией обнаруженной интенсивности, при предположении, что детектор работает ниже предела насыщения19:

В случае фотонововых детекторов 1и с0, таким образом, дисперсия детектора равна средней интенсивности:

Чтобы применить эти понятия к реальным измерениям в живых клетках, Gratton и его коллеги18 определяют кажущуюся яркость, B, для каждого пикселя как соотношение дисперсии над средней интенсивностью:

B является параметр, который измеряется экспериментально. В этой работе, временные ряды изображения рецепторов FGFR1 на плазменной мембране клеток HeLa захвачены tIRF микроскопии и средняя кажущаяся яркость, B, определяется анализом N и B. Затем, после добавления FGF2, последовательные временные ряды фиксируются, чтобы следить за изменениями в самосборке молекул рецепторов на поверхности мембраны после стимуляции рецептора каноническим лигандой.
Однако, поскольку детектором микроскопа TIRF является камера EMCCD, выражение видимой яркости должно быть изменено как19:

где смещение является смещение интенсивности обнаружения электроники, что является характерной для настроек детектора. Дисперсия и средняя интенсивность для аналогового детектора, соответственно, даются:
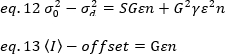
где G является аналоговым увеличением в цифровых уровнях (DL/фотоны), S, цифровые уровни на фотон19, дается наклон интенсивности против дисперсии участка для источника света с постоянной интенсивностью (без временных колебаний). Коэффициент q связан с формой объема обнаружения пикселей. По данным Hassler et al.32, коэффициент q равен 0,3 для tIRF изображения, работающего при максимальном выигрыше камеры обнаружения19. Параметры смещения, S и G являются характеристиками камеры и микроскопа. Очевидная яркость, B, получена путем перестановки eq. 11 в соответствии с eq. 12 и 13:

Экспериментально, является сложной функцией интенсивности лазера и эффективности обнаружения системы. Тем не менее, поскольку B/S линейно зависит от К, важно определить только относительную стоимость к для данного режима обнаружения:

где « пропорционально К. Тем не менее, калибровка выполняется с помощью внутренней ссылки.
