Describimos un enfoque de imagen de Fluorescencia-Número de Fluorescencia Y Brillo de Reflexión Interna Total (TIRF-N&B) para determinar el estado oligomérico promedio de las moléculas receptoras en la membrana plasmática de las células vivas, con el objetivo de vincular el conjunto del receptor dinámica a la función biológica de las proteínas (Figura 1).
Tras la unión de ligandoextra extracelular, los receptores inician la transducción de la señal intracelular dependiendo de su conformación, oligomerización, posibles coreceptores y composición de la membrana. A pesar de la importancia y ubicuidad de la oligomerización de receptores, reconocido como un evento clave en la señalización celular1,2,3,4,5,6, 7, pocos métodos pueden detectar eventos de agrupación en clústeres y medir el grado de agrupación en clústeres experimentalmente8,9. El volumen confocal (x,y a 300 nm, z a 900 nm) no está suficientemente resuelto para probar la interacción molecular y la estequiometría, incluso después de la optimización por algoritmos de restauración de imágenes10. La composición de la subunidad de los oligómeros proteicos no puede resolverse sobre una base puramente espacial incluso mediante métodos de superresolución a resolución x,y de 20-70 nm como PALM11,STORM12y STED13. Además, su resolución temporal (en el orden de minutos por imagen) no puede seguir la cinética en el rango de segundos. El blanqueamiento escalonado de una sola molécula resuelve la estequiometría de los oligómeros proteicos sólo si están inmóviles14.
Uno de los métodos más versátiles para medir la densidad y el oligomerización de proteínas etiquetadas fluorescentes dentro de imágenes individuales es el análisis de distribución de intensidad espacial (SpIDA), que se basa en el muestreo espacial. Es aplicable tanto a células químicamente fijas como vivas, y permite el análisis de varias regiones de interés de la célula simultáneamente utilizando la microscopía de fluorescencia estándar15. Alternativamente, los métodos de momento, tales como espectroscopia de fluorescencia-correlación (FCS)16, histograma de recuento de fotones (PCH)17,y Número y brillo (N&B)18,19, son adecuados para el oligomer cuantitativo Medidas. Estos métodos analizan las fluctuaciones de intensidad de fluorescencia que se pueden observar en el tiempo cuando los fluoróforos se difunden dentro y fuera de un volumen de iluminación. Las amplitudes de las fluctuaciones de intensidad pueden describirse de forma única por el brillo molecular del fluoróforo y el número medio de fluoróforos (n) dentro del volumen de iluminación17 (Figura 2). Típicamente, el coeficiente de difusión de los fluoróforos y el número medio de moléculas (inversamente relacionadas con el valor G(0)) dentro del volumen de iluminación se pueden obtener por FCS20. Sin embargo, dado que el tiempo de difusión sólo se escala con la raíz cúbica de la masa, el FCS no es lo suficientemente sensible para detectar cambios en la masa molecular21. En la práctica, fcS de un solo color no puede detectar la dimerización de los receptores de membrana. PCH resuelve mezclas de diferentes oligómeros con precisión. Utilizando más de dos momentos de la distribución de amplitud, detecta moléculas de diferente brillo que ocupan el mismo volumen de iluminación. Escaneando FCS22 y desarrollos, como la interesante correlación par del enfoque23del brillo molecular (pCOMB), introducido para ampliar el rango de aplicabilidad de los métodos de correlación de fluorescencia en los sistemas biológicos24 , siguen siendo métodos de un solo punto que carecen de la capacidad de mediciones rápidas en un área grande de una celda, lo que requiere muchas observaciones consecutivas en cada píxel y la adquisición de datos en el orden de segundos.
N&B es una versión simplificada de PCH que considera sólo el primer y segundo momento de la amplitud de la distribución de la fluorescencia, a saber, la intensidad media, , y la varianza, n.o2 (Figura 2)18,19 y, debido a eso, no puede determinar la fracción molar de los oligómeros desconocidos en una mezcla, pero sólo estima el estado medio de oligomerización de la mezcla. Sin embargo, N&B tiene la ventaja de trabajar con series temporales relativamente más pequeñas de imágenes de células en vivo que PCH en una base de píxel a píxel, simplemente monitoreando las fluctuaciones en el tiempo de la intensidad de la fluorescencia. Debido a que N&B reduce el tiempo por píxel a unos pocos microsegundos, puede seguir la cinética de oligomerización rápida en áreas de celdas grandes, lo que permite la adquisición de imágenes en una escala de tiempo de segundos en microscopía de escaneo ráster (por ejemplo, confocal, 2 fotones) y milisegundos de milisegundos microscopía basada en cámara (por ejemplo, TIRFM).
Varios informes han demostrado la capacidad de N&B para cuantificar el número de subunidades en los grupos de proteínas mediante imágenes de regiones celulares extendidas. Se detectaron grupos de Paxillin-EGFP en los sitios de adhesión en las células CHO-K125,y la agregación intracelular del péptido patógeno Httex1p se describió en las células COS-726. N&B se aplicó para seguir la oligomerización impulsada por ligando sorda del receptor ErbB27, y el efecto del ligando FGF21 en Klothob (KLB) y FGFR1c en las células De LaLa28. La combinación de imágenes TIRF y análisis N&B se utilizó para mostrar que el dinamina-2 es principalmente tetramírico en toda la membrana celular29. Aplicamos N&B tanto a las imágenes de escaneo ráster como a las imágenes TIRF para probar la dimerización impulsada por ligando sin efecto de los receptores de membrana celular uPAR y FGFR130,31.
Los métodos de correlación de fluorescencia, como N&B, FCS y PCH, se basan en la noción de que en un volumen abierto el número de ocupación de partículas sigue una distribución de Poisson. Debido a que sólo se pueden detectar los fotones que emiten los fluoróforos, el valor  medio de una intensidad de fluorescencia medida frente al tiempo en un píxel de la imagen, , es el producto del número medio de fluoróforos en el volumen de iluminación, n, y su brillo molecular,17:
medio de una intensidad de fluorescencia medida frente al tiempo en un píxel de la imagen, , es el producto del número medio de fluoróforos en el volumen de iluminación, n, y su brillo molecular,17:

donde se expresa como el número de fotones emitidos por unidad de tiempo (convencionalmente por segundo) por molécula cuando la molécula está en el centro del volumen de iluminación.
El brillo es una propiedad de cada fluoróforo en una adquisición determinada establecida, mientras que la intensidad es la suma de todas las contribuciones de todos los fluoróforos. En los concursos biológicos, el brillo aumentará con el aumento del número de fluoróforos que fluctúan juntos, dando información sobre el estado de oligomerización de la proteína etiquetada fluorescentemente. Las amplitudes de fluctuación en un píxel dado se miden a partir de la varianza de la señal de fluorescencia,n.o 2:

Donde la media del cuadrado  de intensidad, , y el
de intensidad, , y el  cuadrado de la media de intensidad, , se calculan a partir de los valores de intensidad individuales en cada píxel de cada fotograma:
cuadrado de la media de intensidad, , se calculan a partir de los valores de intensidad individuales en cada píxel de cada fotograma:
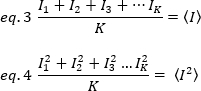
donde K es el número total de fotogramas de la serie temporal. Experimentalmente, es necesario calcular para toda la serie de imágenes la varianza que describe la dispersión de los valores de intensidad individuales en cada píxel de una sola imagen alrededor del valor de intensidad media. La varianza incluye todas las fluctuaciones de diferentes orígenes. En una primera aproximación, la varianza debida a las partículas difusoras en el volumen de iluminación,20, se puede separar de la varianza debido al ruido de disparo del detector,2d. Las dos varianzas son independientes; por lo tanto, la desviación total se da por su suma:

La varianza, debido a las fluctuaciones moleculares dentro y fuera del volumen de detección, depende linealmente del brillo molecular y la intensidad:

Reorganización eq. 6 según eq. 1:

Según el concepto típico en espectroscopía de correlación de fluorescencia, la ecuación 7 establece que la varianza debida al número de fluctuaciones depende del cuadrado del brillo de las partículas.
Entonces, la varianza debida a las fluctuaciones del detector es una función lineal de la intensidad detectada, bajo el supuesto de que el detector se opera por debajo de su límite de saturación19:

En el caso de los detectores de recuento de fotones a unnúmero 1 y ca 0, por lo tanto, la varianza del detector es igual a la intensidad media:

Para aplicar estos conceptos a mediciones reales en celdas vivas, Gratton y sus colegas18 definen el brillo aparente, B, para cada píxel como la proporción de la varianza sobre la intensidad media:

B es el parámetro que se mide experimentalmente. En este trabajo, las imágenes de series temporales de receptores FGFR1 en la membrana plasmática de las células HeLa son capturadas por la microscopía TIRF y el brillo aparente promedio, B, está determinado por el análisis N&B. Luego, después de la adición de FGF2, series temporales consecutivas se capturan para seguir los cambios en el autoensamblaje de las moléculas receptoras en la superficie de la membrana después de la estimulación del receptor con el ligando canónico.
Sin embargo, dado que el detector del microscopio TIRF es una cámara EMCCD, la expresión para el brillo aparente debe modificarse como19:

donde el desplazamiento es el desplazamiento de intensidad de la electrónica de detección que es una característica de la configuración del detector. La varianza y la intensidad media de un detector analógico se dan respectivamente por:
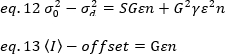
donde G es la ganancia analógica en niveles digitales (DL/fotones), S, los niveles digitales por foton19,se da por la pendiente de una intensidad frente a la varianza de la gráfica para una fuente de luz con intensidad constante (sin fluctuaciones temporales). El factor de la unidad está relacionado con la forma del volumen de detección de píxeles. Según Hassler et al.32, el factor de á es igual a 0,3 para las imágenes TIRF que funcionan con la ganancia máxima de la cámara de detección19. Los parámetros offset, S y G son características de la cámara y del microscopio. El brillo aparente, B, se obtiene reorganizando eq. 11 según eq. 12 y 13:

Experimentalmente, es una función compleja de la intensidad del láser y la eficiencia de detección del sistema. Sin embargo, puesto que B/S depende linealmente de la variable, sólo es importante determinar el valor relativo de la opción para un modo de detección determinado:

donde es proporcional a . Aún así, una calibración se realiza utilizando una referencia interna.
