Biosensor-expressing human pseudoislets were created via the adenoviral delivery of constructs encoding the cAMP biosensor cADDis (Figure 1A). Figure 1B shows the reaggregation of the transduced human islet cells over time, with fully formed pseudoislets observed after 6 days of culture. The cells began to show visible cADDis fluorescence within 48 h, and there was high biosensor expression in transduced cells by the end of the culture period. Using this protocol, the pseudoislets displayed an average transduction efficiency of 60% in alpha cells and 95% in beta cells (Figure 1C–D).
The integrated microfluidic and live-cell imaging platform is highlighted in Figure 2. After harvesting the cADDis-expressing pseudoislets and allowing them to incubate in fresh medium overnight, the pseudoislets were loaded into the center of a pre-wetted well on a microchip. The microchip was then clamped in a holder and placed on a microscope stage. Inside the environmental chamber attached to the microscope, which was maintained at 37 °C, the pseudoislets were perifused with media containing alpha and beta cell secretagogues, which were pumped through a de-bubbler to prevent the introduction of air bubbles into the system. The pseudoislets in the microchip were perifused at a flow rate of 100 µL/min, and the effluent was collected at 2 min intervals via a fraction collector (Figure 2A–B). The glass bottom of the microchip well allows for capturing the cADDis fluorescence intensity of multiple pseudoislets in a field of view, which is essential for later analysis (Figure 2C).
Figure 3 shows representative results using the described protocol with cADDis-expressing pseudoislets generated from native islets isolated from three human donors without diabetes. The cADDis-expressing pseudoislets were perifused with 2 mM glucose (G 2), 20 mM glucose plus the phosphodiesterase inhibitor isobutylmethylxanthine (IBMX; G 20 + IBMX), and 2 mM glucose plus epinephrine (G 2 + Epi). This protocol elicits known intracellular pathways that enhance cAMP within alpha and beta cells (Figure 3A–B). The microperifusion setup facilitates the synchronous collection of the intracellular cAMP dynamics through cADDis fluorescence and hormone secretion (Figure 3C–E). To ensure experimental rigor, experiments on the pseudoislets were performed from the same donor with two to three replicates, and the aggregate values are plotted in Figure 3C–E,D',E'. With exposure to G 2, the relative cADDis intensity was low and stable, as was the insulin secretion (Figure 3C–D). When the pseudoislets were exposed to G 20 + IBMX, there was a robust increase in the intracellular cAMP concentration, as evidenced by an increase in the relative cADDis intensity (Figure 3C). This increase was most likely due to both beta and alpha cells, as demonstrated by the concomitant marked increases in both insulin and glucagon secretion (Figure 3D–E). The exposure of the pseudoislets to G 2 + Epi increased the intracellular cAMP concentration (Figure 3C), and this was associated with an increase in glucagon secretion (Figure 3E). The observation that the cAMP response to G 2 + Epi was relatively smaller compared to the IBMX response may have occurred due to a combination of the lower transduction efficiency of the cADDis adenoviral construct in alpha cells versus beta cells (Figure 1D) and the alpha cell-specificity of this signal (Figure 3E). Therefore, based on known cAMP-mediated signaling pathways in primary human beta and alpha cells, this protocol successfully demonstrates the utility of this integrated platform to simultaneously examine the relative fluorescence intensity of the cADDis biosensor and beta and alpha cell hormone secretory profiles across technical and biological replicates.
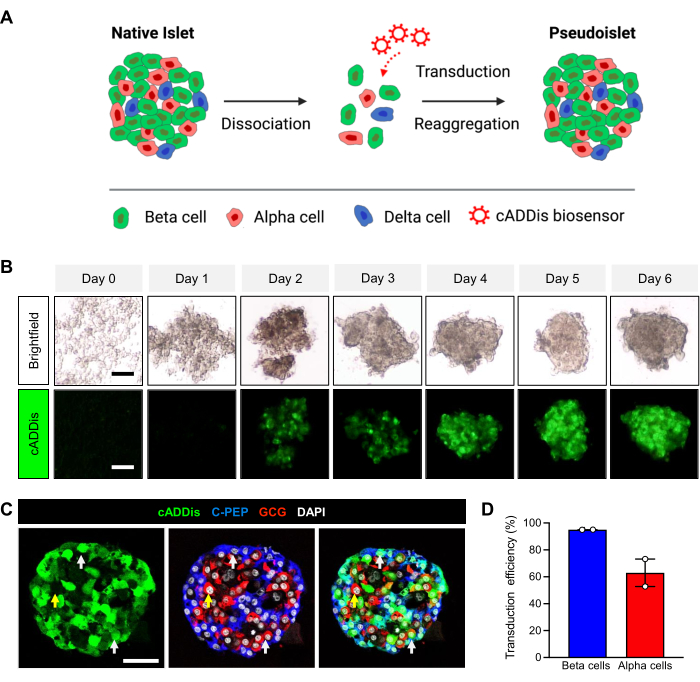
Figure 1: Formation of biosensor-expressing human pseudoislets. (A) Schematic showing primary human islet dissociation, transduction with the biosensor construct in a single-cell state, and reaggregation. Following reaggregation, the pseudoislets are harvested and undergo synchronous live-cell imaging and microperifusion. The protocol steps are indicated throughout the schematic. (B) Representative images capturing the formation of a cADDis-expressing pseudoislet during a 6 day reaggregation period; top: brightfield with contrast, bottom: cADDis fluorescence. The scale bar is 100 µm. (C) Representative images of a cADDis-expressing pseudoislet (green) with alpha and beta cells visualized by C-peptide (CPEP, blue) and glucagon (GCG, red) labeling, respectively. DAPI (white) was used as a nuclear counterstain. The white arrows highlight the transduced alpha and beta cells. The yellow arrow points to an untransduced alpha cell. The scale bar is 100 µm. (D) Quantification of the transduction efficiency at MOI 1,000 in beta and alpha cells by a HALO cytonuclear algorithm (N = 2 donors, with an average of 601 beta cells and 627 alpha cells analyzed per donor across multiple pseudoislets). The error bars indicate the standard error of the mean. Please click here to view a larger version of this figure.

Figure 2: Integrated live-cell imaging and microperifusion system. (A) Overview of the live-cell imaging and microperifusion system components. The pseudoislets in a microchip are placed on the stage of a confocal laser-scanning microscope enclosed within an environmental chamber. This configuration allows for temporal resolution regarding the intracellular changes in biosensor fluorescence during the dynamic response to a series of well-defined secretagogues and the collection of pseudoislet perifusate fractions for the synchronous measurement of the hormone secretion for further integration with intracellular signaling events. (B) The components of the microfluidic platform outside of the environmental chamber. The fluid directionality is as follows: (1) secretagogue-containing tubes to (2) 0.01 in tubing to (3) peristaltic pump tubing (0.02 in inner diameter) to (4) 0.01 in tubing to (5) de-bubbler to (6) microchip inlet (0.01 in tubing) to (7) microchip holder/microchip to (8) microchip outlet (0.01 in tubing). Note: The 0.01 in tubing is joined to the peristaltic pump tubing with conical adapters (white arrowheads). The tubing is plugged into the de-bubbler via a nut and ferrule (green arrowhead). The dashed red line displays the connectivity between the components. (C) Close-up view of the microchip and holder within the environmental chamber mounted on the confocal microscope stage. Please click here to view a larger version of this figure.
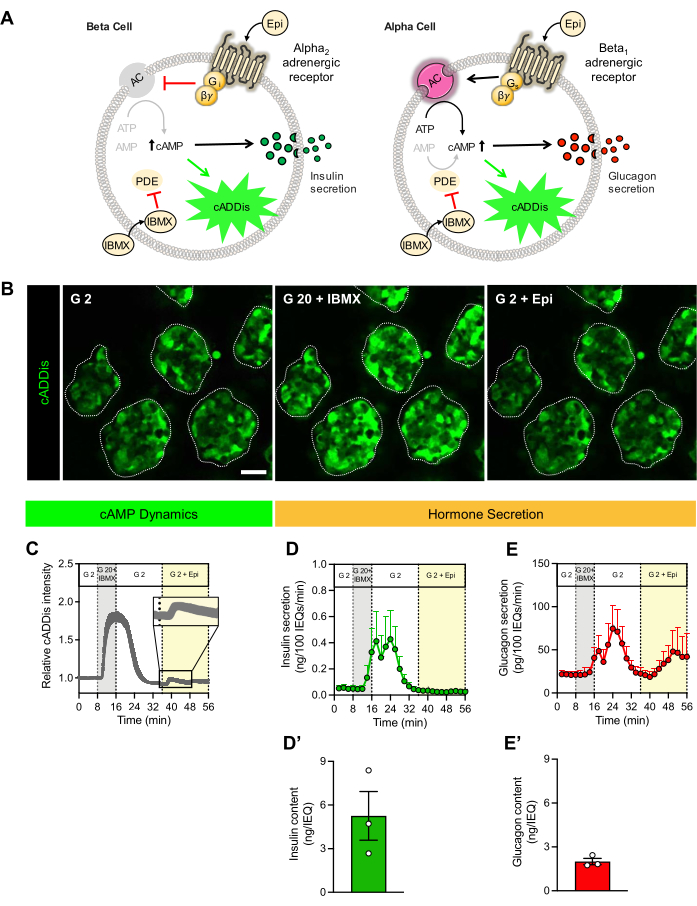
Figure 3: Human pseudoislet cAMP and hormone secretion dynamics. (A) Schematics of secretagogue-induced cADDis fluorescence and hormone secretion in beta and alpha cells. In beta cells, epinephrine (Epi) binds Gi-coupled alpha2-adrenergic receptors, which leads to the inhibition of adenylyl cyclase (AC), decreased cAMP, and reduced insulin secretion. In alpha cells, Epi stimulates Gs-coupled beta1-adrenergic receptors, leading to the activation of AC and an increase in intracellular cAMP. In both cell types, IBMX, a phosphodiesterase (PDE) inhibitor, prevents the degradation of intracellular cAMP and promotes hormone secretion. The binding of free intracellular cAMP causes conformational changes in the cADDis protein, thus causing an increase in fluorescence intensity. (B) Representative images of cADDis-expressing pseudoislets during live-cell imaging in response to G 2 (2 mM glucose), G 20 + IBMX (20 mM glucose + IBMX), and G 2 + Epi (2 mM glucose + Epi). The pseudoislets are outlined in white, and the secretagogue type is indicated in the top-left corner. The scale bar is 50 µm. (C) Relative cADDis fluorescence over time averaged across cADDis-expressing pseudoislets made from human islets from three organ donor preparations. (D) Aggregate insulin and (E) glucagon secretion in response to the indicated secretagogues. The data are normalized to the islet volume, expressed in islet equivalents (IEQs). The pseudoislets were prepared from three human islet donors and analyzed using two to three technical replicates/donor. The islet cells were transduced with cADDis adenoviral construct at MOI 500 (N = 1) or 1,000 (N = 2). (D',E') The pseudoislet hormone content. The error bars indicate the standard error of the mean. Please click here to view a larger version of this figure.
Table 1: Donor demographics. Summary of donor demographic information for the human islets used during the preparation of the pseudoislets with cADDis biosensor expression and live-cell imaging. Please click here to download this Table.
Table 2: Troubleshooting guide. Challenges that may be encountered are highlighted, including a list of common causes of issues, solutions, and prevention techniques. Please click here to download this Table.
Table 3: Microperifusion protocol. An example microperifusion protocol used for the experiments highlighted in the representative results. This protocol is specifically geared toward the co-registration of intracellular cAMP and islet hormone secretion; alternate protocols may be more appropriate depending on the intracellular molecule of interest and/or the chosen biosensor dynamics. Please click here to download this Table.
Supplementary File 1: The calculation of islet equivalents (IEQ). An example of how to calculate the IEQ for the pseudoislets used in each experiment. The data in Sheet 1 (1. Object Measurements) include object data exported directly from the software (see step 9.1). The green highlighted column is inserted to count the number of pseudoislets that fall within each index in step 9.1.6.2. The counts are copied to Sheet 2 (2. IEQ Calculation) and converted to the IEQ using the appropriate conversion factor per index. Any pseudoislet loss during chip loading (In-chip pseudoislet #) and prior to the removal of extracts (Post-run pseudoislet #) can be accounted for using the images captured during these phases (see step 5.5 and step 7.1.1, respectively). Please click here to download this File.
Supplementary File 2: Data analysis. An example of how to calculate the relative cADDis intensity from the data obtained in step 9.2. The raw data in columns D-F is exported directly from the software. The raw values at each time point are normalized to the average intensity in the last minute of baseline medium (G 2; 7-8 min in the protocol). The average intensity across all the pseudoislets over time is calculated to generate the graph in Figure 2C. Please click here to download this File.
