When preparing CFAI media, glucose was exchanged for an increase in lactose and glycerol as the main energy substrate in the media. Additionally, the buffering capacity of the CFAI media was increased as well. These specific components are given in Table 1.
The cells were then grown to both an OD600 of 10 and the standard 2.5 in CFAI media to show consistency with extract quality despite varying extract quantities. The 2.5 OD600 CFAI media was grown after inoculating from a seed culture in LB broth at 37 °C, 200 rpm, while the OD600 10 culture was inoculated directly from a plate. Each batch of CFAI media was then monitored and harvested at their respective OD600. The growth to an OD600 of 10 led to an increase in higher amount of cell pellet and overall extract obtained, as it produced 9.60 mL of extract versus the 2.10 mL of extract obtained from the growth to 2.5 OD600 (Figure 2). Further analysis of total protein concentration demonstrated no significant difference in overall protein in each extract (Table 5). Even though they were grown to different levels of optical density, both batches of extract demonstrated similar results in cell-free reactions using sfGFP (Figure 3). This suggests that the combination of the increased buffering capacity, the use of lactose and glycerol as the main carbon source and implementing lactose instead of IPTG for T7RNAP induction help stabilize extract growths to any OD600 below 10.
| Autoclaved CFAI Media: | |
| Components | Amount |
| Sodium Chloride | 5.0 g |
| Tryptone | 20.0 g |
| Yeast Extract | 5.0 g |
| Potassium Phosphate, monobasic | 6.0 g |
| Potassium Phosphate, dibasic | 14.0 g |
| NanopureTM Water | Fill to a total of 960 mL |
| Filter Sterilized Sugar Solution: | |
| Components | Amount |
| D-Glucose | 0.50 g |
| D-Lactose | 4.0 g |
| 80% v/v Glycerol | 7.5 mL |
| NanopureTM Water | 28.0 mL |
Table 1: CFAI components. Components for CFAI media and sugar solutions with their respective amounts. The media should be stirred throughout the addition of each component and the sugar solution filter sterilized. Each solution should be added to a separate sterile container prior to inoculation.
| S30 Buffer | |
| Components | Concentration |
| Tris Acetate pH 8.2 at room temperature | 10 mM |
| Magnesium Acetate | 14 mM |
| Potassium Acetate | 60 mM |
| Dithiothreitol | 2 mM |
Table 2: S30 buffer components: Components for S30 buffer were added with their respective amounts into a sterile 50 mL conical tube.
| Component | Amount |
| Solution A | 2.20 μL |
| Solution B | 2.1 μL |
| Extract | 5 μL |
| DNA Template | Volume for 16 μg/mL final |
| Water | Fill to a total of 15 μL |
Table 3: CFPS reaction ratios: Relative volume percentages for Solution A, Solution B, and extract. The DNA volume can vary depending on the specific plasmid's concentration and may need to be optimized for the user's specific plasmid being used.
| Solution A | Solution B | ||
| Components | Concentration | Components | Concentration |
| ATP | 1.2 mM | Magnesium Glutamate | 10 mM |
| GTP | 0.850 mM | Ammonium Glutamate | 10 mM |
| UTP | 0.850 mM | Potassium Glutamate | 130 mM |
| CTP | 0.850 mM | Phosphoenolpyruvate (PEP) | 30 mM |
| Folinic Acid | 31.50 μg/mL | L-Valine | 2 mM |
| tRNA | 170.60 μg/mL | L-Tryptophan | 2 mM |
| Nicotinamide Adenine Dinucleotide (NAD) | 0.40 mM | L-Isoleucine | 2 mM |
| Coenzyme A | 0.27 mM | L-Leucine | 2 mM |
| Oxalic Acid | 4.00 mM | L-Cysteine | 2 mM |
| Putrescine | 1.00 mM | L-Methionine | 2 mM |
| Spermidine | 1.50 mM | L-Alanine | 2 mM |
| HEPES Buffer pH 7.5 | 57.33 mM | L-Arginine | 2 mM |
| L-Asparagine | 2 mM | ||
| L-Aspartic Acid | 2 mM | ||
| L-Glutamic acid | 2 mM | ||
| L-Glycine | 2 mM | ||
| L-Glutamine | 2 mM | ||
| L-Histidine | 2 mM | ||
| L-Lysine | 2 mM | ||
| L-Proline | 2 mM | ||
| L-Serine | 2 mM | ||
| L-Threonine | 2 mM | ||
| L-Phenylalanine | 2 mM | ||
| L-Tyrosine | 2 mM | ||
Table 4: Solution A and B components. Stock concentrations for the components for Solution A and B were added with their respective amounts, each in a 1.5 mL microfuge tube.
| Extract | Total Protein Concentration (μg/mL) | Standard Deviation |
| 2xYTPG 2.5 OD | 30617 | 3745 |
| CFAI 2.5 OD | 30895 | 2254 |
| CFAI 10.0 OD | 27905 | 3582 |
Table 5: Total extract protein yields. Analysis of the total protein of different cell extract growths. Total protein concentration was determined using a Bradford Assay. Each concentration was determined from triplicates using a 1:40 dilution.
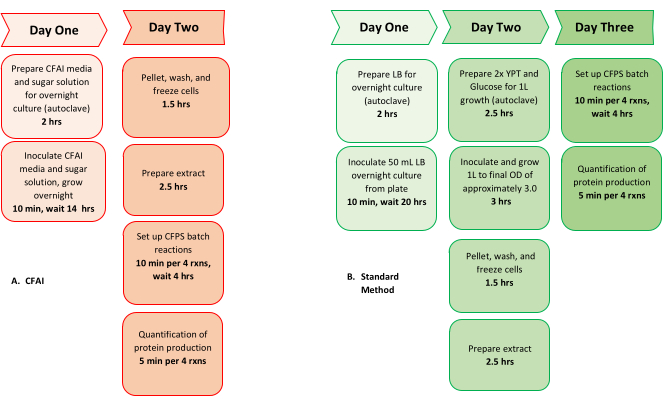
Figure 1: Comparison of CFAI and typical workflow from cells to CFPS: Comparison of the overall timeline from cells to CFPS using the (A) CFAI workflow (left, red) versus the (B) previously established method (green, right). The comparison demonstrates the reduced researcher oversight and timeline when performing CFPS using the CFAI workflow. Please click here to view a larger version of this figure.

Figure 2: CFAI pellet size comparison. Comparison of CFAI media pellets after cell harvest at different OD600. The media grown to an OD600 of 2.5 produced a 2.23 g cell pellet (left) and the media grown to an OD600 of 10 produced a 9.49 g cell pellet (right). Please click here to view a larger version of this figure.
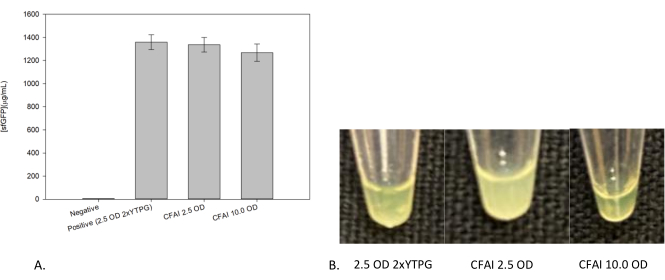
Figure 3: Effects of growth on CFPS reaction yields. (A) Comparison of CFPS reaction yields between growths to 2.5 OD600 and 10 OD600, with (B) images of each CFPS reaction above their respective yield. Cell-free reactions were performed in a 1.5 mL microfuge tube and quantified after 24 h of incubation at 37 °C using a standard curve to correlate fluorescence to sfGFP concentration. The "Negative" corresponds to the set of negative control reactions in which no template DNA was added. The traditional 2xYTPG media (the positive control) and the CFAI extracts are of similar quality as demonstrated through their high CFPS yields. Please click here to view a larger version of this figure.
