単細胞受精卵から有機体の基本的なボディ計画を開発します。ほとんどの顕花植物のココヤシ部門は、樹状突起と撮影やルート、1それぞれに発展する基底のセルを生成します。したがって、胚発生に伴う植物体を形成する方法を理解することが重要だが、彼らは花に深く成長するのでリビング受精および胚のダイナミクスを直接観測する効果的なツールをされていません。トウモロコシ、米などのいくつかの単子葉種体外受精法は確立された2,3をされています。この方法では分離精子と卵細胞を電気的または化学的に、融合し、生成されたセル、肥沃な工場に開発。ただし、双子葉植物植物の雄性と雌性の配偶子の4、5の細胞周期の非同期状態のためおそらく適切な胚を作り出すことができる体外受精法ではありません。さらに、胚周囲組織 (胚乳) は、胚の開発6で重要な役割を果たしています。
モデルの双子葉植物の種、シロイヌナズナ、体外の栽培方法は、胚と胚乳の7を含む全体の胚珠に焦点を当てによって開発されました。このシステムは正常に様々 な化学試薬の胚発生に及ぼすを分析に使用されたが、低い生存率をもっているため時間経過イメージ投射のため適していません。そこで、新規培養胚珠の栽培システムは、受精卵の段階として早期に開始高比8で肥沃な植物を生成するために開発されました。様々 な試験後 Nitsch 培地が分かったし、トレハロースが胚珠8の生存率を大幅に向上します。さらに、胚珠の展開としては成長して、従って頻繁に顕微鏡の観察フィールドから移動、ために、9の中の胚珠を修正する PDMS 装置が開発されました。PDMS デバイスは、心期胚には受精卵から開発をトレースするのに十分である 3-4 日間の長期的なイメージングが有効になります。このメソッドを使用して、それは偏波受精卵・胚のパターンだけでなく通常の条件下でも低分子阻害剤の存在下でまたは様々 な突然変異体の背景8,10 のダイナミクスを可視化することが可能になります。 ,11。
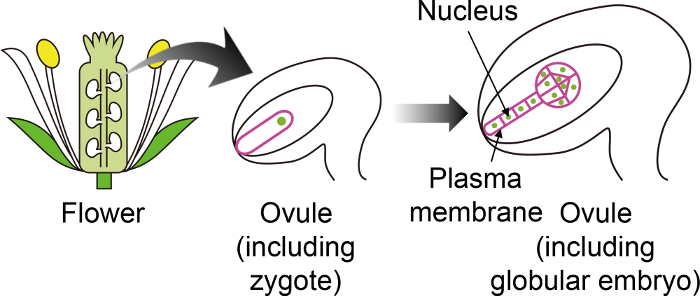
図 1: 視覚化受精し胚を胚珠を使用特定の蛍光マーカーの模式図。
シロイヌナズナの受精卵は、花の奥深くに生成される胚珠の胚に開発しています。体外培養システム、受精卵・胚胚珠、観察、従って他の胚珠組織で表されない特定の蛍光マーカーを使用することが重要です。 この図の拡大版を表示するのにはここをクリックしてください。
