בשני העשורים האחרונים, האסטרטגיה כתב כימי התפתחה מתודולוגיה שני שלבים חזק כדי לחקור את הדינמיקה ואת הפונקציות של מולקולות שאינן גנטית מקודד. 1 , 2 , 3 באסטרטגיה אנלוגי סינתטי של biomolecule עניין עם אפנון קטן – כתב כימי – עובר תחילה מטבוליזם האורגניזם החי, ולאחר מכן בדיקה כימית (למשל., fluorophore על ידי קרינה פלואורסצנטית מיקרוסקופיה קונפוקלית הדמיה) קשורה covalently הכתב incorporated ויה bioorthogonal לחץ על כימיה. המכשיר חייב להגיב במהירות, במיוחד עם השינוי כימי הציג בעת היותו אינרטי כדי כל מולקולות נוכח במערכת החיים. במובנים רבים, שיטה זו גוברת על המגבלות של טכניקות נפוצות bioconjugation באמצעות לחץ ספציפי מאוד כימיה ligations ובכך מספק הזדמנות לעקוב מטבוליטים או ובמקרו-מולקולות ביולוגיות כי היו נגישים בעבר חי מערכות4,5,6.
למרות הפופולריות בצמיחה מהירה של שיטה זו עוצמה של תאים חיידקיים ובעלי חיים, דוחות המתאר את השימוש בביולוגית הצמח הם כמה מפתיע, המרחק בין7,8,9,10, 11,12. היינו מעוניינים במיוחד ליישם אסטרטגיה זו בצמחים ללמוד היווצרות של ליגנין, לאחד biopolymers הנפוץ ביותר על פני כדור הארץ, מרכיב עיקרי של ביומסה lignocellulosic. 13 , 14 ליגנין הוא פולימר בעל תאים פנוליים זה ממלא תפקיד חיוני הגנה מבניים בתחום הפיתוח החיים של צמחים.
הוא בדרך כלל מורכב moieties 4-hydroxyphenylpropanoid שלושה: H (p– hydroxyphenyl), G (guaiacyl) ויחידות S (syringyl) נגזר בהתאמה בין 3 ‘monolignols’ (p– coumaryl, coniferyl ו- sinapyl כהלים) כי הם מסונתז דרך השביל phenylpropanoid בציטופלסמה של התא (איור 1). לאחר להיות מיוצא דופן התא, monolignols הם מחומצן לקיצוניים peroxidases או laccases לאחר מכן הם עוברים צימוד הרדיקלי גרידא כימי תגובות פולימריזציה כדי ליגנין פולימרים, תהליך הנקרא lignification. 15 , 16 למרות lignins חריפה להשפיע על valorization תעשייתי של רבים הצמחי מוצרים, הקהילה המדעית יש עדיין דרך ארוכה ללכת כדי לפענח את המנגנון הסבוך ויסות lignification לו.
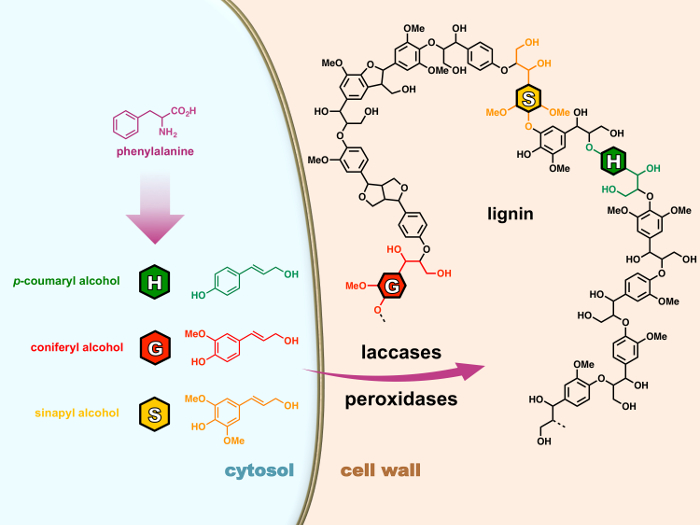
איור 1: תהליך lignification בתאי צמח. Monolignols הם biosynthesized של פנילאלנין ב ציטוזול. לאחר להיות מיוצא דופן התא, monolignols הם מחומצן לקיצוניים peroxidases או laccases לאחר מכן הם עוברים צימוד הרדיקלי גרידא כימי תגובות פולימריזציה כדי ליגנין פולימרים, תהליך הנקרא lignification. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
למרות דיווחים על השימוש bioorthogonal תגובות לניתוח glycan רבים,2,3,17 שלהם יישום דוגמאות לסוגים אחרים של מולקולות הן פחות. השימוש bioorthogonal כימיה למטרות bioimaging ליגנין רק לאחרונה היה חלוץ ידי. Tobimatsu et al. 8 ב תודרנית לבנה כדי לספק מידע אודות שילוב של coniferyl המחליפים אלכוהול לתוך הפולימר ליגנין איפה זה יוצר את יחידות G,8,9 ובכך הממחיש את הוכחת את זה כתב כימי אסטרטגיות ישימות בהקשר זה. השימוש CuAAC היה ללמוד גם באמצעות נגזרת אלכוהול שונים coniferyl חודש כמה מאוחר יותר בוקובסקי ואח. 9 . אולם, ליגנין מכיל גם יחידות H ו- S הבנה עמוקה יותר של תהליך lignification דורש יותר ידע איך כל monolignols משולבים הפולימר, מה הגורמים עשויות לשלוט הקומפוזיציה שלה. הפיתוחים החדשים בתחום זה כיום תלויים פיתוח מתודולוגיות יעיל כדי לעקוב אחר כתבים כימיות מרובות בו-זמנית במערכות החיים. אף-על-פי כמה כתבות על glycans הניחו את היסודות בשנים האחרונות18,19,20,21,22, כפול תיוג גישות נשארים אתגר גדול ב bioorthogonal כימיה. אם פרוטוקול לחץ לשחזור תיוג-יחיד הוא קשה לפתח, ואז כפול תיוג גישות הדורשים אופטימיזציה במשולב שני הדדית תואמת bioorthogonal תגובות על שני הכתבים כימיים נפרדים הם אפילו יותר קשה. כמה דוגמאות כי חלוץ היבט זה השתמשו בשילוב של אזיד-אלקין אתקרב זן cycloaddition (SPAAC) ותגובות תגובת דילס-אלדר (DAinv) דרישה אלקטרונית הופכי אלקן-tetrazine ללמוד glycans בתאים בעלי חיים. עם זאת, חשבנו כי bioorthogonality התגובה DAinv לא יכול להיות מובטח ביישום זה בשל המאפיינים המבניים של ליגנין (אשר מורכב של מונומרים אלקטרון-עשיר cinnamyl שהוחלפו מסוג זה יכול להגיב עם אלקטרון עלוב האוויר דינס כגון הגששים tetrazine המשמש בתגובות DAinv), כי זה עלול לייצר שאינם ספציפיים תיוג. בנוסף, התגובהinv DA דורש ידיות כימיים קשים לאכסון גישה, כמו גם להיות מגושם ולא lipophilic ובכך מעלים את האפשרות כי הקצב של התאגדות, תחבורה ו/או לוקליזציה של החומר הכימי כתב ויוו עלולים להיות מושפעים. כפי שקלנו כי ההיבט האחרון היה רלוונטי במיוחד במקרה של גישה כימיה בלחיצה ללמוד lignification, אנחנו בחרה בכיוון שונה ופיתח באמצעות Bioorthogonal מצדו הדמיה רציפים אסטרטגיה (אושר) שילוב של Cycloaddition אזיד-אלקין Strain-Promoted (SPAAC) ו- Cycloaddition אזיד-אלקין קטליזת של נחושת (CuAAC) ויוו. 23
תגובות שני אלו הם אכן שני bioorthogonal הראשי לחץ על תגובות בהן השתמשו עד כה, יותר בפרט כמה דוגמאות של ליגנין הדמיה אשר פורסמו לאחרונה. 8 , 9 אסטרטגיית תיוג כפול שלנו מאפשרת את השימוש של אזיד moiety על monolignol אחד כתב וידיות של אלקין מסוף מצד שני, שני הכימי i) אינרטיים לכיוון מבנים רלוונטי מבחינה ביולוגית, ii) מאוד קטנים בגודלם (איור 2 ). כתוצאה, ההשפעה של אלה שינויים סינתטי על המאפיינים physicochemical של biomolecule שנבחנה ממוזער ובכך להקטין אפשרי אי-התאמות בין סובסטרטים לא טבעיים, הטבעית monolignol מבחינת תחבורה, metabolization המחירים במהלך השלב התאגדות מטבולית. למרות השילוב של SPAAC ו- CuAAC נראה מאוד אינטואיטיבי במבט ראשון, זה לידע שלנו רק הדוגמה השניה של סימון כפול באמצעות אסטרטגיה זו לבין היישום הראשון על מבנים שאינם glycans. 12 , 23
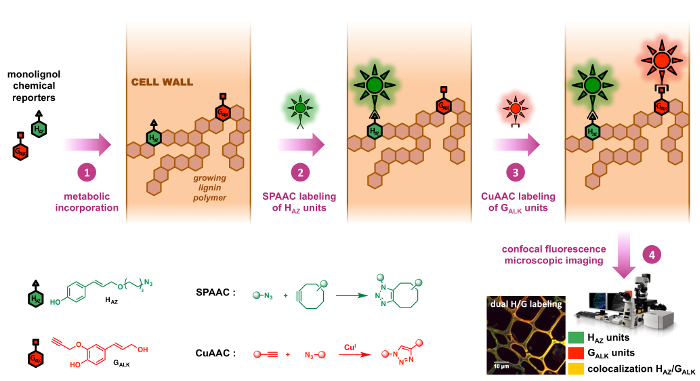
איור 2: אושר כפול תיוג אסטרטגיה. כתבים כימי HAZ GALK מתויג אנלוגים של monolignols מקורית H ו- G בקרב אנשי עסקים ותיירים כאחד. הם משולבים קודם פולימרים ליגנין הגוברת של קירות התא על ידי האכלת אקסוגני (שלב 1). Cyclooctyne – ו אזיד-functionalized הגששים פלורסנט ואז ברצף מאתרים בפני הכתבים incorporated מאת bioorthogonal לחץ על כימיה: התגובה SPAAC (שלב 2) הוא מאוד ספציפי של יחידות HAZ , ואחריו (התגובה CuAAC שלב 3) זה מפורט של GALK יחידות (שלב 3), ובכך לאפשר לוקליזציה ספציפי של שני עיתונאים באופן עצמאי במדגם זהה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
אנו ראשית המיועד לאמת את monolignol מתויג אזיד כתב HAZ (ממלא מקום של p– coumaryl אלכוהול), קודמן של יחידות ליגנין H ו ואז המציאו את אושר כפול תיוג האסטרטגיה שבה הוא משמש במשולב עם קודם לכן דווח מתויג אלקין GALK,9 (ממלא מקום של אלכוהול coniferyl), קודמן של ליגנין G יחידות. ב פרוטוקול לשחזור זה פיתח ובחן, פשתן, זן צמח חשוב מבחינה כלכלית, שילוב מטבולית כפול של HAZ ו- GALK לתוך ליגנין מושגת קודם לפני רציפים SPAAC/CuAAC תיוג. כאן, מתויג HAZ יחידות תחילה במפורש מסומנות באמצעות מצדו SPAAC של fluorophore cyclooctyne-functionalized, ואחריו בתיווך CuAAC מצדו של בדיקה פלורסנט השני במתויג GALK יחידות. שיטה זו שימש לחקור את הדינמיקה של תהליכים lignification בתוך קירות התא צמח וניתן יישומית ויוו לבלום חתכים, חיים נובע, כמו גם אחרים-שתילים, מינים שונים.
