過去 20 年間の化学薬品レポーター戦略動態および非遺伝子にエンコードされた生体分子の機能を調査するための強力な 2 段階の方法論として浮上しています。1,2,3この戦略で小さな変調-化学薬品レポーター -興味の生体分子の合成アナログはまず代謝される生きている有機体の化学プローブ順 (e.g、蛍光の蛍光。共焦点顕微鏡イメージング) 共有、bioorthogonal クリックケミストリーによって組み込まれた記者にリンクされます。急速に、特にプローブが反応する必要があります任意の生体分子の生きているシステムに存在する不活性でありながら導入化学修飾。多くの方法でこのメソッドは、非常に特定のクリック化学およびそれにより代謝物または以前アクセスできなかった生体高分子を追跡する機会を提供するを使用して共通化反応技術の限界を克服します。生活システム4,5,6。
細菌と動物の細胞でこの強力なメソッドの急成長している人気にもかかわらず植物生物学での使用を記述するレポートは、意外にも少数ありまでの間7,8,9,10、します。 11,12。地球上最も一般的な生体高分子の一つ、リグノ セルロース系バイオマスの主成分リグニンの形成を研究する植物でこの戦略を適用することに特に興味を持っていた。13,14リグニンはフェノール樹脂開発と高等植物の生活保護構造の重要な役割を果たしています。
一般的に 3 つの 4 hydroxyphenylpropanoid 鎖から成り立っています: H (p-ヒドロキシフェニル)、G (グアヤシル) と S (結果) の単位はそれぞれ 3 つから派生した、’monolignols’ (p– coumaryl、コニフェリルとシナピル アルコール)フェニルプロパノイド経路 (図 1) 細胞の細胞質で合成されました。Monolignols の酸化細胞壁にエクスポートされている後ペルオキシダーゼまたはリグニン ポリマー重合する純粋化学の根本的なカップリング反応を受けるその後 laccases によってラジカルにプロセスは木質化と呼ばれます。15,16リグニンに強く影響を与える植物由来の多くの産業の物価安定政策が、製品、科学界は、木質化を調節する複雑なメカニズムを解読する長い道のりをまだ持っています。
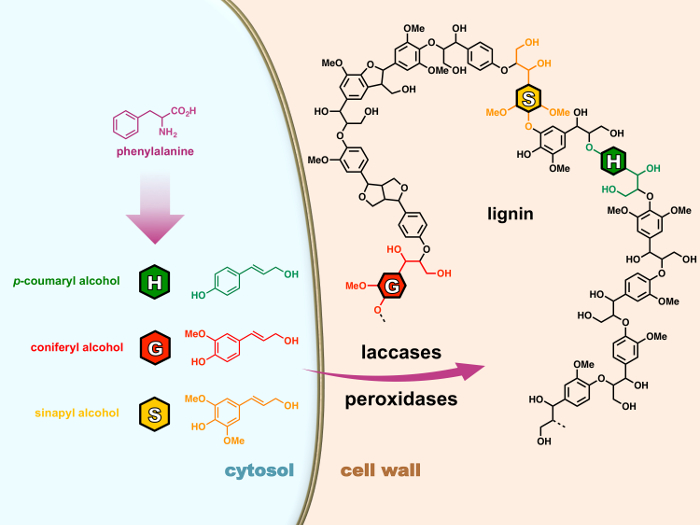
図 1: 植物細胞における木質化プロセス。Monolignols は細胞質でフェニルアラニンから生合成されます。Monolignols の酸化細胞壁にエクスポートされている後ペルオキシダーゼまたはリグニン ポリマー重合する純粋化学の根本的なカップリング反応を受けるその後 laccases によってラジカルにプロセスは木質化と呼ばれます。この図の拡大版を表示するのにはここをクリックしてください。
糖鎖解析 bioorthogonal 反応の利用に関する報告が多数、2,3,17他生体分子の種類にその応用例は少ない。リグニン バイオ イメージング用 bioorthogonal 化学の使用が飛松らによって開拓されたばかり8 シロイヌナズナG 単位、8,9概念実証デモを形作るそれリグニン ポリマーにアルコール サロゲート コニフェリル定款についての情報を提供するために、化学薬品レポーターの戦略は、このコンテキストで適用されます。CuAAC の使用もによって示された数ヶ月別のコニフェリル アルコール誘導体を使用して後でブコウスキーら。9ただし、リグニンも H と S のユニットを含むし、木質化のプロセスのより深い理解は、monolignols のすべてが、ポリマーに組み込まれる方法についてより多くの知識を必要とするどのような要因がその組成を制御します。このフィールドの新しい進歩は現在生きているシステムで同時に複数の化学記者を追跡するための効率的な手法の開発に依存します。主要な課題にもかかわらず、糖鎖に関するいくつかの記事は、近年18,19,の基礎を築いたが20,21,22, デュアル アプローチをラベル付けのままbioorthogonal 化学。再現可能な単一ラベル クリック プロトコルが開発するは難しい、互いに 2 つのタンデムで最適化を必要とするアプローチをラベリングし、デュアルの場合 2 つの独立した化学記者に互換性のある bioorthogonal 反応がさらに困難。この面を開拓したほとんどの例は、動物細胞の糖鎖を勉強するひずみ昇格アジ化物アルキン環 (SPAAC) とアルケン tetrazine 逆の電子式需要 (DAinv) のディールス ・ アルダー反応の組み合わせを しました。ただし、我々 は、DAinv 反応の bioorthogonality は、(電子不足と反応することができます電子豊富な置換シンナミル タイプ単量体から成っているリグニンの構造機能のおかげでこのアプリケーションの保証されないかもしれないと思ったジエン DAinv 反作用で使用された tetrazine のプローブなど)、これが非固有のラベルを生成する可能性があります。加えて、DAinv反応がかさばるし、可能性を高めることにより脂溶性にあることと同様、アクセス困難な総合的化学処理が必要ですが設立、輸送および/または化学物質の局在化の率生体内で記者があります。後者の側面は木質化を研究するためのクリック化学アプローチの場合特に関連すると考えて、我々 別の方向を選んだ、Bioorthogonal 結紮イメージング逐次戦略 (至福) を使用して開発、Strain-Promoted アジ化物アルキン環 (SPAAC) と銅触媒によるアジ化物アルキン環 (CuAAC) の組み合わせ体内。23
これら 2 つの反応は、実際に 2 つの主な bioorthogonal をクリックして日付に使用されている反応とリグニンのいくつかの例では特に画像最近出版されました。8,9デュアル ラベリング戦略を有効に 1 つモノリグノール記者アジ化部位と、他のターミナル アルキン i) 関連する生物学的構造にたがわず、ii) は非常に小さなサイズ (図 2の 2 つの化学ハンドルの使用).その結果、調査の下で生体分子の物理化学的性質のこれらの合成の修正の影響を最小限交通の面で不自然なモノリグノール基板間不一致を減らすと代謝率代謝設立段階。SPAAC と CuAAC の組み合わせが、一見非常に直感的なようだがデュアルのマーキング以外の糖鎖構造にこの作戦の最初のアプリケーションを使用してのみ、2 番目の例我々 の知識です。12,23
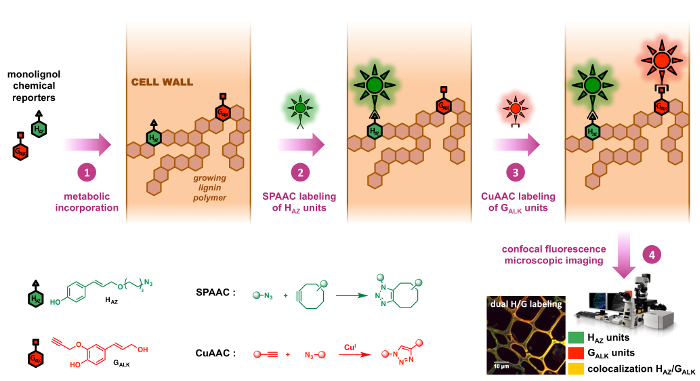
図 2: ブリス デュアル戦略をラベリングします。化学記者 HAZ GALKは、ネイティブの H と G monolignols の類似するタグ。彼らは最初外因性 (ステップ 1) を供給することにより、細胞壁の成長のリグニンの重合体に組み込まれます。Cyclooctyne とアジ化-機能集積化蛍光プローブが、順番に組み合わされて組み込まれた記者に bioorthogonal によってクリックケミストリー: SPAAC 反応 (ステップ 2) 特異性が高く Hアリゾナ州単位の CuAAC 反応 (が続きますステップ 3) 単位 GALK (ステップ 3) の特定であるようにない両方の記者の特定のローカリゼーション独立して同じサンプルの。この図の拡大版を表示するのにはここをクリックしてください。
我々 は、まず設計アジ化タグ モノリグノール記者HAZ ( p– coumaryl アルコールの代理人) とリグニン H 単位の前駆体を検証し、連動される至福のデュアル ラベリング戦略を考案、タグ付きのアルキンGALK9 (コニフェリル アルコールの代理人) とリグニン G 単位の前駆体を報告しました。この再現可能なプロトコルを開発し、亜麻のテストでは、経済的に重要な植物、リグニンにHAZとGALKのデュアル代謝定款はシーケンシャル SPAAC/CuAAC の前に達成した最初ラベリング。ここでは、タグHアリゾナ州単位最初具体的にはラベルが付いています経由でタグGALK の 2 番目の蛍光プローブの CuAAC を介した結紮に続いて cyclooctyne 修飾 fluorophore の SPAAC 結紮ユニット。このメソッドは、植物細胞壁内の木質化のプロセスのダイナミクスを調べるに使用された、応用生体内で茎の断面、別の植物種の苗と同様、茎の生活をすることができます。
