Det tværpolitiske GAL4-UAS-system er arbejdshesten for funktionel karakterisering af gener i insektmodelorganismen Drosophila melanogaster1,2,3. For at bruge GAL4-UAS-systemet krydses transgene førerlinjer, der udtrykker gærtransskriptionsfaktoren GAL4 under kontrol af en lovgivningsmæssig sekvens, med responderlinjer, der bærer et interessegen eller RNA-interferens (RNAi) konstruktion kontrolleret af en Upstream Activation Sequence (UAS), der er anerkendt af GAL4. Afkommet af dette kors udtrykker transgene af interesse i en spatiotemporal mønster dikteret af promotor kontrollerende GAL4 udtryk (figur 1). Fænotyper, der vises af afkom af driver-responder kors kan vurderes at belyse funktionen af kandidat gener. Selvom D. melanogaster er blevet brugt til at undersøge gener fra andre organismer4,5,6,7, er GAL4-UAS-systemet nu blevet tilpasset til brug i insekter af medicinsk og landbrugsmæssig betydning for at give direkte analyse af interessearter 8,9,10,11,12,13,14.
I den afrikanske malaria myg, Anopheles gambiae, GAL4-UAS-systemet blev først testet af celle linje co-transfection9. Flere konstruktioner blev analyseret for effektivitet i forskellige parvis kombinationer og fandt, at 14 tandemly gentagne UAS suppleret med en lille kunstig intron (UAS-14i) viste det bredeste udvalg af aktiveringspotentiale, når de anvendes med et panel af GAL4 drivere. For at demonstrere in vivo-funktionalitet blev disse konstruktioner derefter brugt til at skabe to separate transgene An. gambiae linjer af PiggyBac transformation8: en førerlinje, der transporterer GAL4 drevet af en midgutspecifik promotor, og en responderlinje, der indeholder både luciferase og forbedrede gule fluorescerende protein (eYFP) gener under regulering af UAS-sekvenser. Tarmspecifik luciferaseaktivitet og fluorescens i afkommet indikerede, at systemet var effektivt i Anopheles. Siden da er der blevet oprettet driverlinjer, der udtrykker transgener i andet væv, der er vigtigt for vektorkapacitet og insekticidresistens, herunder oenocytter15 og hæmocytter16, og i et tæt på allestedsnærværende mønster10. Talrige UAS linjer er også blevet genereret til analyse gener menes at være involveret i stofskiftet og beslaglæggelse medieret insekticidresistens, cuticular kulbrinte syntese og fluorescerende tag forskellige celle-og vævstyper (tabel 1). For responder linjer, site-rettet integration af transgene er nu udført af ΦC31 katalyseret rekombination kassette udveksling17,18 at fastsætte den genomiske sammenhæng af UAS regulerede gener. På denne måde normaliseres transgene udtryk med hensyn til genomisk indsættelsessted, hvilket giver mulighed for en mere præcis sammenligning af de fænotypiske virkninger af forskellige kandidatgener.
De responder linjer skabt til dato er designet til enten at udtrykke transgene enten på forhøjede niveauer eller til at reducere genekspression gennem RNA interferens (RNAi). Normalt cDNA kloner er smeltet til UAS sekvens til at generere egnede udtryk plasmider, men fuld genomiske sekvenser er også muligt under forudsætning af, at de ikke er for store til kloning. For at generere lyddæmpningskonstruktioner har vi brugt tre forskellige metoder til at opnå egnede tandem omvendte sekvenser, der danner hårnål dsRNA, der stimulerer RNAi. Disse har inkluderet fusion PCR, asymmetrisk PCR og kommerciel syntese af hårnål konstruktioner. Fælles for hver metode er inkluderingen af en intronsekvens mellem de omvendte sekvenser for at give kloningsstabilitet. Responder plasmider, hvori et gen af interesse / RNAi konstruktion kan indsættes er blevet udviklet15. Disse plasmider bærer også de krævede ΦC31 attB-steder til RMCE (beskrevet i Adolfi, der ledsager JoVE-papir, der beskriver RCME-teknikken i detaljer). Protokoller, der dækker de vigtige trin, der kræves, når du vælger sekvensen til indsættelse i en af disse plasmider til overekspression, er inkluderet i dette manuskript. Derudover er to protokoller for RNAi hårnål konstruere skabelse beskrevet og illustreret.
Når du opretter nye linjer, er identifikation af sjældne transgene individer afgørende at opdrætte fra at etablere og opretholde transgene kolonier. Vigtigst for GAL4-UAS-systemet er det nødvendigt at skelne mellem responder- og førerlinjerne for at etablere kryds og identificere individuelle afkom, der bærer begge transgener. Dette opnås ved at bruge forskellige dominerende valgbare markørgener knyttet til driveren og responderkassetter. Oftest er disse fluorescerende markørgener, der klart kan skelnes ved hjælp af optiske filtre (f.eks. eYFP, eCFP, dsRed). Det er vigtigt, at markører udtrykkes i et kendt og pålideligt spatiotemporalt mønster, da dette gør identifikation af abnormiteter og forurening lettere. Fluorescerende markør genekspression er rutinemæssigt reguleret af den syntetiske 3xP3 promotor, som forårsager øje og ventrale ganglier specifikke udtryk i alle faser af An. gambiae udvikling19. Fluorescerende markører kontrolleret af 3xP3 er inkluderet i alle transformation plasmider beskrevet i denne artikel. En protokol, der beskriver de almindelige metoder, der anvendes til at screene fluorescerende An. gambiae pupae GAL4-UAS linjer er inkluderet her.
Et af nøgleelementerne i GAL4-UAS-systemet er nødvendigheden af at krydse de differentierede driver- og responderlinjer. For at gøre dette skal mandlige og kvindelige fra hver linje adskilles før parring. Voksne er let skelnes ved synet, men for at etablere genetiske kors er det fornuftigt at adskille kønnene før voksen fremkomst for at sikre, at parring ikke har fundet sted. Den generelle størrelse forskel mellem mandlige og kvindelige An. gambiae pupper er for variabel til at være en effektiv og pålidelig metode til kønsbestemmelse20. I stedet giver klare morfologiske forskelle i de ydre kønsorganer et pålideligt grundlag for sexing i An. gambiae. I denne artikel beskriver vi en pålidelig metode til sexing An. gambiae pupper til at oprette passende kors.
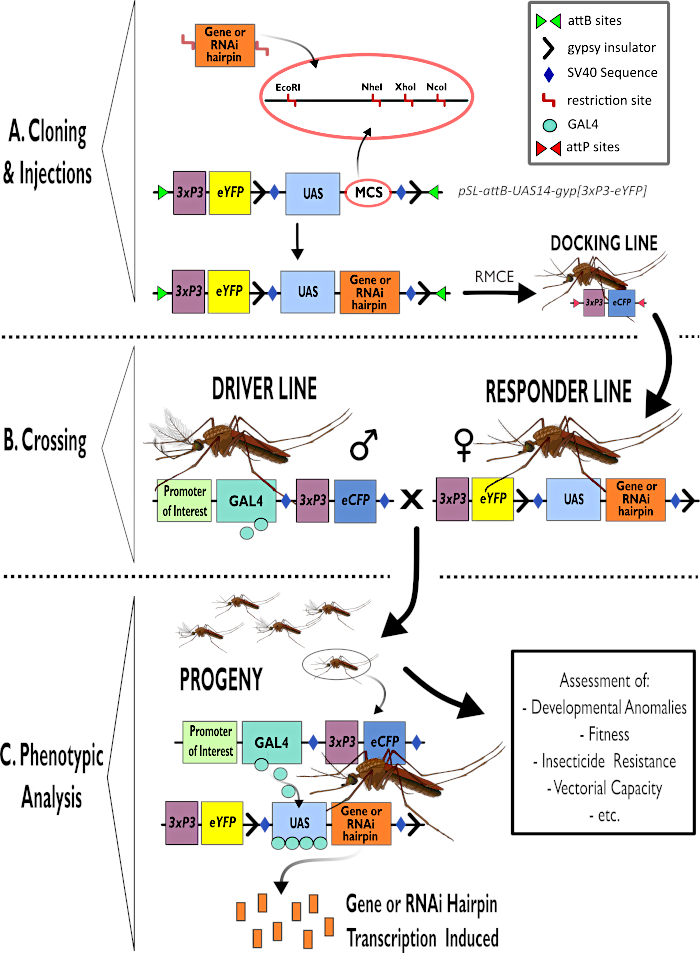
Figur 1 – Diagrammatisk repræsentation af processen for anvendelse af det todelte GAL4-UAS-system i Anopheles gambiae. (A) De vigtigste komponenter i en eksempelvektor (pSL-attB-UAS14-gyp[3xp3-eYFP]) er afbildet med detaljer om de tilgængelige begrænsningssteder (EcoRI, NheI, XhoI og NcoI) inden for de flere kloningssteder, der er egnede til brug til at indsætte hårnålkonstruktions- eller kodningssekvensen for interessegen. Dockinglinjens struktur er også afbildet. (B) Overgangstrinnet illustreres med angivelse af mænds brug fra førerlinjen (med GAL4-driver af en initiativtager af interesse og eCFP drevet af 3xP3-promotoren) og kvinder fra responderlinjen (med interessegen eller hårnålkonstruktionen kontrolleret af en UAS-promotor og en eYFP-markør, der kontrolleres af 3xP3-promotoren). (C) En diagrammatisk gengivelse af GAL4-kørselsekspression af det interessegen, der er af interesse for korsets afkom i B, og en liste over nogle af de typiske fænotyper, der vurderes. Forkortelser: Multiple Cloning Site (MCS), Recombinase medieret kassetteudveksling (RMCE), Upstream Activator Sequence (UAS), forbedret gult fluorescerende protein (eYFP), forbedret cyan fluorescerende protein (eCFP). Klik her for at se en større version af dette tal.
Det er brugen af kryds, der giver gal4-UAS-systemets todelte karakter, som har klare fordele i forhold til mere lineære tilgange. F.eks. kan mange flere kombinationer af fører- og reaktionslinjer vurderes, end det ville være muligt, hvis der skulle genereres og opretholdes en ny transgen linje for hver promotor/genkombination. Endnu vigtigere, det giver mulighed for analyse af gener, der producerer dødelige eller sterile fænotyper, når deres udtryk er forstyrret, som er vanskelige at skabe / vedligeholde i et lineært system. Sådanne dødelige fænotyper kan manifestere sig på alle udviklingsstadier, afhængigt af genfunktionen og spatiotemporal ekspression, men observeres oftest under embryonal udvikling. Visualisering myg embryo udvikling kræver clearing af uigennemsigtig chorion, som frakker æggene. Følgende metoder beskrevet i Trpiš (1970)21 og Kaiser et al. (2014)22 beskriver vi de protokoller, vi bruger til at rette embryoner, samtidig med at vi opretholder strukturel integritet og blegning for at rydde den enkercheion, der tillader mikroskopisk visualisering og billeddannelse.
