מערכת GAL4-UAS הדו-מפלגתית היא סוס העבודה של אפיון תפקודי של גנים באורגניזם מודל החרקים Drosophila melanogaster1,2,3. כדי להשתמש במערכת GAL4-UAS, קווי נהג מהונדסים, המבטאים את גורם שעתוק השמרים GAL4 תחת שליטה על רצף רגולטורי, נחצים עם קווי מגיב הנושאים מבנה של עניין או הפרעות RNA (RNAi) הנשלט על ידי רצף הפעלה במעלה הזרם (UAS) המוכר על ידי GAL4. צאצאי הצלב הזה מבטאים את הטרנסג’ן של העניין בתבנית מרחבית המוכתבת על ידי האמרגן השולט בביטוי GAL4 (איור 1). פנוטיפים המוצגים על ידי צאצאים של צלבים מגיבים לנהג ניתן להעריך כדי להדגיש את הפונקציה של גנים מועמד. למרות D. melanogaster שימש כדי לבחון גנים של אורגניזמים אחרים4,5,6,7, מערכת GAL4-UAS הותאמה כעת לשימוש חרקים בעלי חשיבות רפואית וחקלאית כדי לספק ניתוח ישיר במינים מעניינים 8,9,10,11,12,13,14.
יתוש מלריה אפריקאי, אנופלס גמביה, מערכת GAL4-UAS נבדקה לראשונה על ידי קו התא co-transfection9. מבנים מרובים נידונו ליעילות בשילובים שונים של זוג ומצאו כי 14 UAS חוזרים על עצמם יחד בתוספת intron מלאכותי קטן (UAS-14i) הציג את הטווח הרחב ביותר של פוטנציאל הפעלה בעת שימוש עם לוח של מנהלי התקנים GAL4. כדי להדגים פונקציונליות vivo, מבנים אלה שימשו אז כדי ליצור שני קווי A. gambiae מהונדסים נפרדים על ידי PiggyBac טרנספורמציה8: קו נהג הנושא GAL4 מונע על ידי מקדם ספציפי midgut, וקו מגיב המכיל הן את לוציפראז וחלבון פלואורסצנטי צהוב משופר (eYFP) גנים תחת ויסות רצפי UAS. פעילות לוציפראז ספציפית במעיים פלואורסצנטיות בצאצאים הצביעו על כך שהמערכת יעילה אנופלס. מאז, קווי נהג נוצרו המבטאים טרנסגנים ברקמות אחרות החשובות לקיבולת וקטורית ועמידות לקוטלי חרקים, כולל oenocytes15 ו hemocytes16, ובתבנית קרובה לכל מקום10. קווי UAS רבים נוצרו גם כדי לאבחן גנים שנחשבו מעורבים בחילוף החומרים ועמידות קוטלי חרקים מתווכים, סינתזת פחמימנים cuticular ולתייג באופן פלואורסצנטי סוגי תאים ורקמות שונים (טבלה 1). עבור קווי המגיב, שילוב מכוון אתר של transgene מבוצע כעת על ידי ΦC31 מזורז recombination חילופי 17,18 כדי לתקן את ההקשר הגנומי של הגנים המוסדרים UAS. בדרך זו, ביטוי טרנסג’ן מנורמל לגבי מיקום הכניסה הגנומית, ומאפשר השוואה מדויקת יותר של ההשפעות הפנוטיפיות של גנים מועמדים שונים.
קווי המגיב שנוצרו עד כה נועדו לבטא את הטרנסג’ן ברמות גבוהות או להפחית ביטוי גנים באמצעות הפרעות RNA (RNAi). בדרך כלל שיבוטי cDNA מותכים לרצף UAS כדי ליצור פלסמידים ביטוי מתאים, אולם רצפים גנומיים מלאים הם גם אפשריים בהנחה שהם לא גדולים מדי עבור שיבוט. כדי ליצור מבנים השתקה, השתמשנו בשלוש שיטות שונות כדי להשיג רצפים הפוכים טנדם מתאים היוצרים dsRNA סיכת ראש הממריץ RNAi. אלה כללו פי.סי.אר היתוך, PCR אסימטרי וסינתזה מסחרית של מבנים סיכת ראש. המשותף לכל שיטה הוא הכללה של רצף אינטרוני בין הרצפים ההפוכים כדי לספק יציבות שיבוט. פלסמידים מגיבים שבהם גן של עניין / RNAi מבנה פותחו15. פלסמידים אלה נושאים גם את אתרי ה- ΦC31 ATTB הנדרשים עבור RMCE (המתואר באדולפי המלווה נייר JoVE המתאר בפירוט את טכניקת ה- RCME). פרוטוקולים המכסים את השלבים החשובים הנדרשים בעת בחירת הרצף להכנסה לאחד הפלסמידים האלה לביטוי יתר כלולים בכתב יד זה. בנוסף, שני פרוטוקולים ליצירת מבנה סיכת ראש RNAi מתוארים ומאוירים.
בעת יצירת קווים חדשים, זיהוי של אנשים מהונדסים נדירים הוא חיוני להתרבות כדי להקים ולתחזק מושבות מהונדסות. והכי חשוב עבור מערכת GAL4-UAS יש צורך להבחין בין קווי המגיב והנהג כדי ליצור צלבים ולזהות צאצאים בודדים הנושאים את שני הטרנסגנים. זה מושג באמצעות גנים שונים דומיננטיים סמן לבחירה מקושרים לנהג קלטות מגיב. לרוב אלה הם גנים סמן פלואורסצנטי כי הם בבירור להבחין באמצעות מסננים אופטיים (למשל, eYFP, eCFP, dsRed). חשוב כי סמנים באים לידי ביטוי בתבנית spatiotemporal ידוע ואמין כמו זה עושה זיהוי של חריגות וזיהום קל יותר. ביטוי גן סמן פלואורסצנטי מוסדר באופן שגרתי על ידי מקדם 3xP3 סינתטי, אשר גורם לביטוי ספציפי עין וגרעעים גחוניים בכל שלבי פיתוח An. gambiae19. סמנים פלואורסצנטיים הנשלטים על ידי 3xP3 כלולים בכל פלסמידים טרנספורמציה המתוארים במאמר זה. פרוטוקול המפרט את השיטות הנפוצות המשמשות לסינון קווי גלם גלם GAL4-UAS פלואורסצנטי כלול כאן.
אחד המרכיבים העיקריים של מערכת GAL4-UAS הוא הצורך לחצות את קווי הנהג והמגיב המסומנים באופן דיפרנציאלי. כדי לעשות את זה זכר ונקבות מכל שורה חייב להיות מופרד לפני ההזדווגות. מבוגרים ניתנים להבחנה בקלות על ידי ראייה, עם זאת, עבור הקמת צלבים גנטיים זה הגיוני להפריד את המינים לפני הופעת מבוגר כדי להבטיח כי ההזדווגות לא התרחשה. הבדל הגודל הכללי בין גולם א. גמביה זכר לנקבה משתנה מדי מכדי להיות שיטה יעילה ומהימנה לקביעת מין20. במקום זאת הבדלים מורפולוגיים ברורים באיברי המין החיצוניים מספקים בסיס אמין למין ב – An. gambiae. במאמר זה, אנו מתארים שיטה אמינה למין גולם An. gambiae כדי להגדיר צלבים מתאימים.
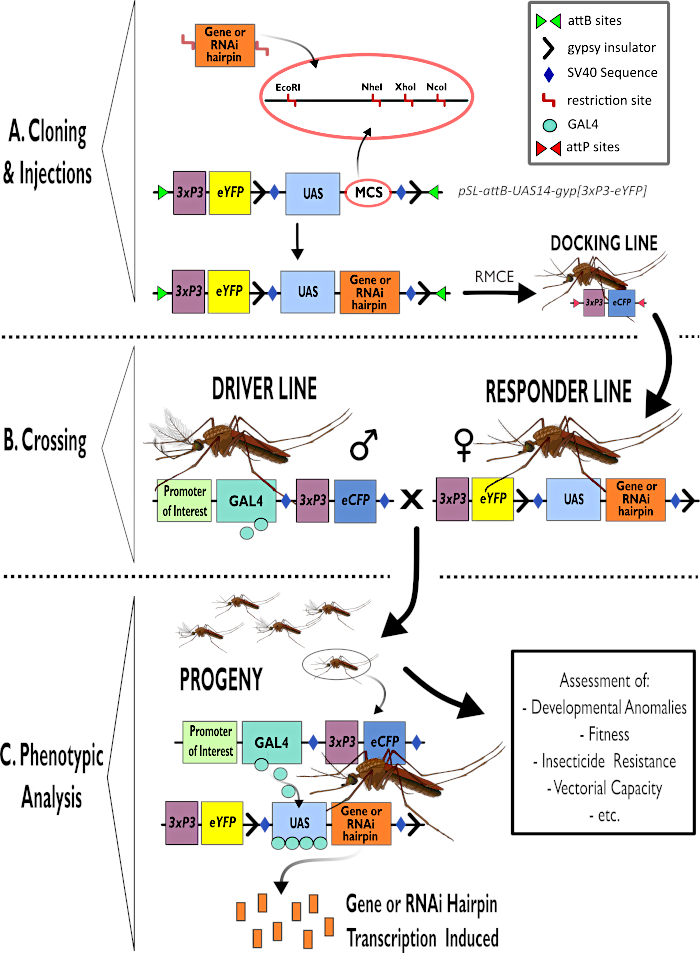
איור 1 – ייצוג דיאגרמתי של תהליך לשימוש במערכת GAL4-UAS הדו-מפלגתית ב-Anopheles gambiae. (A) הרכיבים העיקריים של וקטור לדוגמה (pSL-attB-UAS14-gyp[3xp3-eYFP]) מתוארים, המפרטים את אתרי ההגבלה הזמינים (EcoRI, NheI, XhoI ו- NcoI) בתוך אתרי השיבוט המרובים המתאימים לשימוש כדי להכניס את מבנה סיכת הראש או רצף הקידוד עבור גן העניין. מבנה קו העגינה מתואר גם הוא. (ב) שלב המעבר מומחש המציין את השימוש בזכרים מקו הנהג (נשיאת נהג GAL4 על ידי מקדם עניין ו- eCFP המונע על ידי מקדם 3xP3) ונקבות מקו המגיב (הנושא את גן העניין או מבנה סיכת הראש הנשלט על ידי מקדם UAS וסמן eYFP הנשלט על ידי מקדם 3xP3). (ג) ייצוג דיאגרמתי של ביטוי נהיגה GAL4 של גן העניין בצאצאי הצלב ב- B ורשימה של כמה מהפנוטיפים הטיפוסיים המוערכים. קיצורים: אתר שיבוט מרובים (MCS), החלפת קלטות בתיווך רקומבינאז (RMCE), רצף מפעיל במעלה הזרם (UAS), חלבון פלואורסצנטי צהוב משופר (eYFP), חלבון פלואורסצנטי ציאן משופר (eCFP). אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
זהו השימוש בצלבים המספק את האופי הדו-צדדי של מערכת GAL4-UAS, שיש לה יתרונות ברורים על פני גישות ליניאריות יותר. לדוגמה, ניתן להעריך שילובים רבים יותר של קווי נהג ומגיב מאשר היה אפשרי אם היה צורך ליצור ולתחזק קו מהונדס חדש עבור כל שילוב מקדם / גנים. חשוב מכך, הוא מאפשר ניתוח של גנים המייצרים פנוטיפים קטלניים או סטריליים כאשר הביטוי שלהם מוטרד אשר קשה ליצור / לתחזק במערכת ליניארית. פנוטיפים קטלניים כאלה יכולים להתבטא בכל השלבים ההתפתחותיים, בהתאם לתפקוד הגנים ולביטוי spatiotemporal, אך הם נצפים לרוב במהלך התפתחות עוברית. הדמיית התפתחות העובר יתוש דורש ניקוי של chorion אטום אשר מצפה את הביצים. בעקבות שיטות המתוארות ב- Trpiš (1970)21 וקייזר ואח ‘ (2014)22, אנו מתארים את הפרוטוקולים בהם אנו משתמשים כדי לתקן עוברים, תוך שמירה על שלמות מבנית, והלבנה כדי לנקות את האנדוריון המאפשר הדמיה והפכה מיקרוסקופית.
