Il sistema bipartito GAL4-UAS è il cavallo di battaglia della caratterizzazione funzionale dei geni nell’organismo modello di insetto Drosophila melanogaster1,2,3. Per utilizzare il sistema GAL4-UAS, le linee driver transgeniche, che esprimono il fattore di trascrizione del lievito GAL4 sotto il controllo di una sequenza regolatoria, vengono incrociate con linee di responder portatrici di un costrutto di genico di interesse o di interferenza dell’RNA (RNAi) controllato da una sequenza di attivazione a monte (UAS) riconosciuta da GAL4. La progenie di questa croce esprime il transgene di interesse per un pattern spaziotemporale dettato dal promotore che controlla l’espressione GAL4 (Figura 1). I fenotipi visualizzati dalla progenie di incroci driver-responder possono essere valutati per chiarire la funzione dei geni candidati. Sebbene D. melanogaster sia stato utilizzato per esaminare geni di altri organismi4,5,6,7, il sistema GAL4-UAS è stato ora adattato per l’uso in insetti di importanza medica e agricola per fornire analisi dirette nelle specie di interesse 8,9,10,11,12,13,14.
Nella zanzara africana della malaria, Anopheles gambiae, il sistema GAL4-UAS è stato testato per la prima volta dalla co-trasfezione della linea cellulare9. Sono stati analizzati più costrutti per l’efficienza in diverse combinazioni a coppie e hanno scoperto che 14 UAS ripetuti in tandem integrati con un piccolo introne artificiale (UAS-14i) mostravano la più ampia gamma di potenziale di attivazione quando utilizzati con un pannello di driver GAL4. Per dimostrare la funzionalità in vivo, questi costrutti sono stati poi utilizzati per creare due linee transgeniche Separate an. gambiae mediante la trasformazione piggyBac8: una linea driver che trasporta GAL4 guidata da un promotore specifico del midgut e una linea di risposta contenente sia la luciferasi che i geni della proteina fluorescente gialla potenziata (eYFP) sotto la regolazione delle sequenze UAS. L’attività della luciferasi specifica dell’intestino e la fluorescenza nella progenie indicavano che il sistema era efficiente in Anopheles. Da allora, sono state create linee driver che esprimono transgeni in altri tessuti importanti per la capacità vettoriale e la resistenza agli insetticidi, compresi gli enociti15 e gli emociti16, e in un modello quasi onnipresente10. Sono state inoltre generate numerose linee UAS per analizzare geni ritenuti coinvolti nel metabolismo e nel sequestro mediato dalla resistenza agli insetticidi, dalla sintesi di idrocarburi cuticolari e per etichettare fluorescentemente diversi tipi di cellule e tessuti (Tabella 1). Per le linee di risposta, l’integrazione site-directed del transgene viene ora eseguita mediante scambio di cassette di ricombinazione catalizzate ΦC31111117,18 per fissare il contesto genomico dei geni regolati UAS. In questo modo, l’espressione transgenica viene normalizzata per quanto riguarda la posizione di inserimento genomico, consentendo un confronto più accurato degli effetti fenotipici di diversi geni candidati.
Le linee di risposta create fino ad oggi sono progettate per esprimere il transgene a livelli elevati o per ridurre l’espressione genica attraverso l’interferenza dell’RNA (RNAi). Di solito i cloni di cDNA sono fusi alla sequenza UAS per generare plasmidi di espressione adatti, tuttavia anche le sequenze genomiche complete sono fattibili supponendo che non siano troppo grandi per la clonazione. Per generare costrutti di silenziamento, abbiamo usato tre diversi metodi per ottenere sequenze invertite in tandem adatte che formano dsRNA a forcina che stimola l’RNAi. Questi hanno incluso la PCR di fusione, la PCR asimmetrica e la sintesi commerciale di costrutti a forcina. Comune a ciascun metodo è l’inclusione di una sequenza di introni tra le sequenze invertite per fornire stabilità di clonazione. Sono stati sviluppati plasmidi responder in cui può essere inserito un costrutto gene di interesse/RNAi15. Questi plasmidi portano anche i siti ΦC31 attB richiesti per RMCE (descritti in Adolfi che accompagna l’articolo JoVE che descrive la tecnica RCME in dettaglio). I protocolli che coprono i passaggi importanti richiesti quando si seleziona la sequenza per l’inserimento in uno di questi plasmidi per la sovraespressione sono inclusi in questo manoscritto. Inoltre, vengono descritti e illustrati due protocolli per la creazione di costrutti a forcina RNAi.
Quando si creano nuove linee, l’identificazione di individui transgenici rari è fondamentale per riprodursi per stabilire e mantenere colonie transgeniche. Ancora più importante per il sistema GAL4-UAS c’è la necessità di distinguere le linee di risposta e driver per stabilire incroci e identificare la progenie individuale che trasporta entrambi i transgeni. Ciò si ottiene utilizzando diversi geni marcatori selezionabili dominanti collegati alle cassette del conducente e del risponditore. Più comunemente si tratta di geni marcatori fluorescenti che sono chiaramente distinguibili utilizzando filtri ottici (ad esempio, eYFP, eCFP, dsRed). È importante che i marcatori siano espressi in un modello spaziotemporale noto e affidabile in quanto ciò rende più facile l’identificazione delle anomalie e della contaminazione. L’espressione genica dei marcatori fluorescenti è regolata di routine dal promotore sintetico 3xP3, che causa l’espressione specifica dei gangli oculari e ventrali in tutte le fasi dello sviluppo di An. gambiae19. I marcatori fluorescenti controllati da 3xP3 sono inclusi in tutti i plasmidi di trasformazione descritti in questo articolo. Un protocollo che descrive in dettaglio i metodi comuni utilizzati per schermare le linee fluorescenti An. gambiae pupae GAL4-UAS è incluso qui.
Uno degli elementi chiave del sistema GAL4-UAS è la necessità di attraversare le linee di guida e di risposta con marcatura differenziale. Per fare questo maschio e femmine da ogni linea devono essere separati prima dell’accoppiamento. Gli adulti sono facilmente distinguibili dalla vista, tuttavia, per stabilire incroci genetici è sensato separare i sessi prima dell’emergere degli adulti per garantire che l’accoppiamento non si sia verificato. La differenza generale di dimensioni tra maschio e femmina An. gambiae pupae è troppo variabile per essere un metodo efficiente e affidabile di determinazione del sesso20. Invece chiare differenze morfologiche nei genitali esterni forniscono una base affidabile per il sexing in An. gambiae. In questo articolo, descriviamo un metodo affidabile per il sexing An. gambiae pupae per impostare croci appropriate.
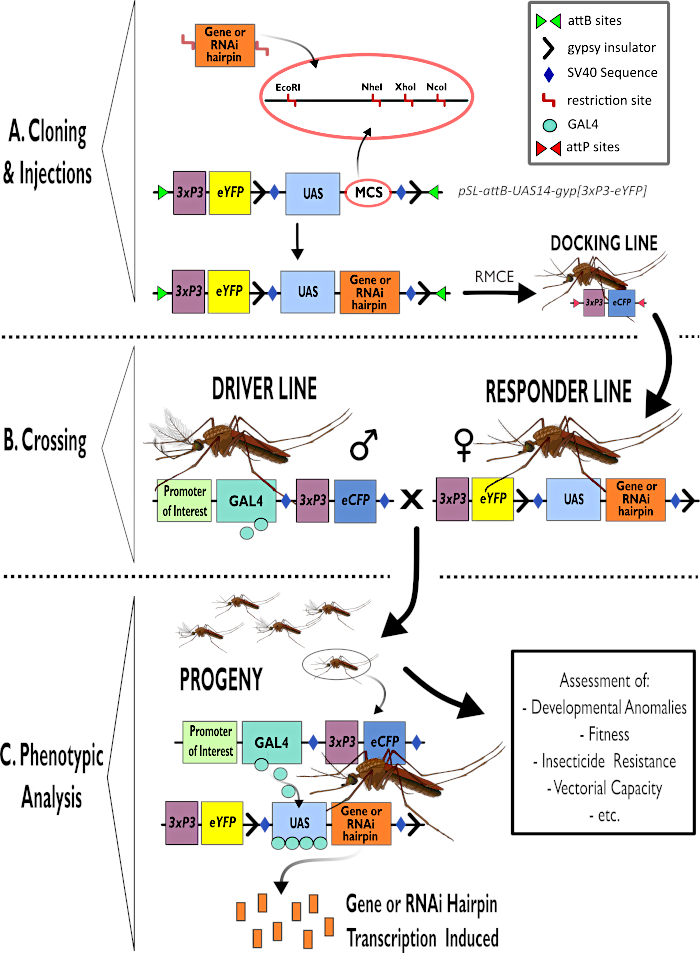
Figura 1 – Rappresentazione diagrammatica del processo per l’utilizzo del sistema bipartito GAL4-UAS in Anopheles gambiae. (A) Sono rappresentati i componenti principali di un vettore di esempio (pSL-attB-UAS14-gyp[3xp3-eYFP]), che descrivono in dettaglio i siti di restrizione disponibili (EcoRI, NheI, XhoI e NcoI) all’interno dei siti di clonazione multipli che sono adatti all’uso per inserire il costrutto a forcina o la sequenza codificante per il gene di interesse. Anche la struttura della linea di attracco è raffigurata. (B) La fase di attraversamento è illustrata indicando l’uso di maschi dalla linea di guida (che trasportano il conducente GAL4 da un promotore di interesse e eCFP guidato dal promotore 3xP3) e femmine dalla linea di risposta (portatori del gene di interesse o del costrutto a forcina controllato da un promotore UAS e da un marcatore eYFP controllato dal promotore 3xP3). (C) Una rappresentazione diagrammatica dell’espressione trainante GAL4 del gene di interesse nella progenie della croce in B e un elenco di alcuni dei fenotipi tipici che vengono valutati. Abbreviazioni: Multiple Cloning Site (MCS), Recombinase mediated cassette exchange (RMCE), Upstream Activator Sequence (UAS), enhanced yellow fluorescent protein (eYFP), enhanced cyan fluorescent protein (eCFP). Fare clic qui per visualizzare una versione più grande di questa figura.
È l’uso delle croci che fornisce la natura bipartita del sistema GAL4-UAS, che presenta vantaggi distinti rispetto agli approcci più lineari. Ad esempio, è possibile valutare molte più combinazioni di linee driver e responder di quanto sarebbe fattibile se una nuova linea transgenica dovesse essere generata e mantenuta per ciascuna combinazione promotore/gene. Ancora più importante, consente l’analisi di geni che producono fenotipi letali o sterili quando la loro espressione è perturbata che sono difficili da creare / mantenere in un sistema lineare. Tali fenotipi letali possono manifestarsi in tutte le fasi dello sviluppo, a seconda della funzione genica e dell’espressione spaziotemporale, ma sono più spesso osservati durante lo sviluppo embrionale. Visualizzare lo sviluppo dell’embrione di zanzara richiede la pulizia del corion opaco che ricopre le uova. Seguendo i metodi descritti in Trpiš (1970)21 e Kaiser et al. (2014)22, descriviamo i protocolli che usiamo per fissare gli embrioni, pur mantenendo l’integrità strutturale, e lo sbiancamento per eliminare l’endocorion che consente la visualizzazione microscopica e l’imaging.
