Bipartit GAL4-UAS sistemi, böcek modeli organizma Drosophila melanogaster1,2,3’teki genlerin fonksiyonel karakterizasyonunun çalışma atıdır. GAL4-UAS sistemini kullanmak için, bir düzenleyici sıranın kontrolü altındaki maya transkripsiyon faktörü GAL4’ü ifade eden transgenik sürücü hatları, GAL4 tarafından tanınan bir Yukarı Akış Aktivasyon Sırası (UAS) tarafından kontrol edilen bir ilgi geni veya RNA paraziti (RNAi) yapısı taşıyan yanıtlayıcı çizgilerle geçilir. Bu haçın soyu, GAL4 ekspresyonunun kontrolüne geçen organizatör tarafından dikte edilen mekansal bir desene olan ilginin transgenesini ifade eder (Şekil 1). Sürücü-yanıtlayıcı haçların soyu tarafından görüntülenen fenotipler, aday genlerin işlevini aydınlatacak şekilde değerlendirilebilir. D. melanogaster diğer organizmalardan gelen genleri incelemek için kullanılmış olmasına rağmen4,5,6,7, GAL4-UAS sistemi şimdi ilgi türlerinde doğrudan analiz sağlamak için tıbbi ve tarımsal öneme sahip böceklerde kullanılmak üzere uyarlanmıştır 8,9,10,11,12,13,14.
Afrika sıtma sivrisinek, Anopheles gambiae, GAL4-UAS sistemi ilk hücre hattı ortak transfection9 tarafından test edildi. Farklı çift yönlü kombinasyonlarda verimlilik için birden fazla yapı test edildi ve küçük bir yapay intron (UAS-14i) ile desteklenmiş 14 tandemly tekrarlanan UAS’ın GAL4 sürücülerinden oluşan bir panelle kullanıldığında en geniş aktivasyon potansiyelini gösterdiği bulundu. In vivo işlevselliğini göstermek için, bu yapılar daha sonra PiggyBac dönüşümü8 tarafından iki ayrı transgenik An. gambiae hattı oluşturmak için kullanıldı: ortagut spesifik bir promotör tarafından sürülen GAL4 taşıyan bir sürücü hattı ve UAS dizilerinin düzenlenmesi altında hem luciferaz hem de gelişmiş sarı floresan protein (eYFP) genlerini içeren bir yanıtlayıcı hattı. Bağırsak spesifik luciferaz aktivitesi ve soydaki floresan, sistemin Anopheles’te verimli olduğunu gösterdi. O zamandan beri, oenositler15 ve hemositler16 da dahil olmak üzere vektörel kapasite ve insektisit direnci için önemli olan diğer dokularda ve her yerde bulunan bir desene yakın bir desende transgenes ifade eden sürücü çizgileri oluşturulmuştur10. Metabolizma ve sequestrasyon aracılı insektisit direnci, kütiküler hidrokarbon sentezi ve farklı hücre ve doku tiplerini floresan olarak etiketlemede rol aldığı düşünülen genleri test etmek için de çok sayıda UAS hattı oluşturulmuştur (Tablo 1). Yanıtlayıcı hatlar için, transjenin raya yönelik entegrasyonu, UAS tarafından düzenlenen genlerin genomik bağlamını düzeltmek için ΦC31 katalze rekombinasyon kaseti değişimi17,18 tarafından gerçekleştirilir. Bu şekilde, transgene ekspresyonu genomik ekleme konumu ile ilgili olarak normalleştirilerek farklı aday genlerin fenotipik etkilerinin daha doğru karşılaştırılmasına olanak sağlar.
Bugüne kadar oluşturulan yanıtlayıcı çizgiler, transjeni yüksek seviyelerde ifade etmek veya RNA paraziti (RNAi) yoluyla gen ekspresyonını azaltmak için tasarlanmıştır. Genellikle cDNA klonları uygun ekspresyon plazmidleri oluşturmak için UAS dizisine kaynaşır, ancak tam genomik diziler klonlama için çok büyük olmadıkları varsayılarak da mümkündür. Susturma yapıları oluşturmak için, RNAi’yi uyaran saç tokası dsRNA’yı oluşturan uygun tandem ters diziler elde etmek için üç farklı yöntem kullandık. Bunlar arasında füzyon PCR, asimetrik PCR ve saç tokası yapılarının ticari sentezi yer alıp almamaktadır. Her yöntem için ortak, klonlama kararlılığı sağlamak için ters diziler arasında bir intron dizisinin dahil edilmesidir. İlgi/RNAi yapısı geninin yerleştirilebileceği yanıtlayıcı plazmidler geliştirilmiştir15. Bu plazmidler ayrıca RMCE için gerekli ΦC31 attB sitelerini de taşır (RCME tekniğini ayrıntılı olarak açıklayan Adolfi eşlik eden JoVE kağıdında açıklanmıştır). Bu makaleye, aşırı ifade için bu plazmidlerden birine ekleme sırası seçilirken gereken önemli adımları kapsayan protokoller yer almaktadır. Ayrıca, RNAi saç tokası yapı oluşturma için iki protokol açıklanmış ve gösterilmiştir.
Yeni çizgiler oluştururken, nadir transgenik bireylerin tanımlanması, transgenik koloniler kurmak ve sürdürmek için üremek için çok önemlidir. Gal4-UAS sistemi için en önemlisi, çaprazlar oluşturmak ve her iki transjeni taşıyan bireysel soyları tanımlamak için yanıtlayıcı ve sürücü hatlarını ayırt etmek bir gerekliliktir. Bu, sürücü ve yanıtlayıcı kasetlere bağlı farklı baskın seçilebilir işaretleyici genleri kullanılarak elde edilir. En yaygın olarak bunlar optik filtreler (örneğin, eYFP, eCFP, dsRed) kullanılarak açıkça ayırt edilebilen floresan işaret genleridir. Anormalliklerin ve kontaminasyonun tanımlanmasının kolay hale getirdiğinden, belirteçlerin bilinen ve güvenilir bir mekansal desende ifade olması önemlidir. Floresan belirteç gen ekspresyumu, An. gambiae gelişiminin tüm aşamalarında göz ve ventral gangliyon spesifik ifadeye neden olan sentetik 3xP3 promotörü tarafından rutin olarak düzenlenir19. 3xP3 tarafından kontrol edilen floresan belirteçler, bu makalede açıklanan tüm dönüşüm plazmidlerine dahildir. Floresan An. gambiae pupa gal4-UAS hatlarını taramak için kullanılan yaygın yöntemleri ayrıntılı olarak açıklayan bir protokol burada yer almaktadır.
GAL4-UAS sisteminin temel unsurlarından biri, farklı şekilde işaretlenmiş sürücü ve yanıtlayıcı hatlarını geçme zorunluluğudur. Bunu yapmak için her satırdan erkek ve dişi çiftleşmeden önce ayrılmalıdır. Yetişkinler göz göre göre kolayca ayırt edilebilir, ancak genetik haçlar oluşturmak için çiftleşmenin gerçekleşmemesini sağlamak için yetişkin ortaya çıkmasından önce cinsiyetleri ayırmak mantıklıdır. Erkek ve dişi An. gambiae pupa arasındaki genel boyut farkı, etkili ve güvenilir bir cinsiyet belirleme yöntemi olamayacak kadar değişkendir20. Bunun yerine dış cinsel organdaki açık morfolojik farklılıklar , An. gambiae’de seks için güvenilir bir temel sağlar. Bu yazıda, uygun haçları kurmak için An. gambiae pupaları seks yapmak için güvenilir bir yöntem açıklıyoruz.
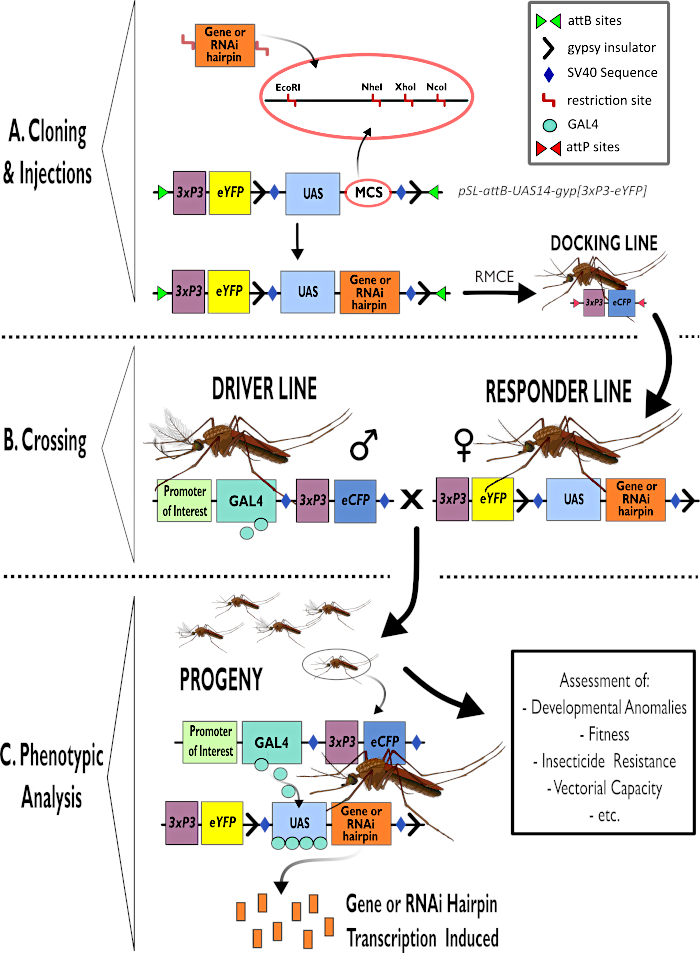
Şekil 1 – Anopheles gambiae’de bipartit GAL4-UAS Sisteminin kullanımına yönelik sürecin şemamatik gösterimi. (A) Örnek bir vektörün ana bileşenleri (pSL-attB-UAS14-gyp[3xp3-eYFP]), ilgi çekici gen için saç tokası yapısı veya kodlama sırası eklemek için kullanıma uygun çoklu klonlama siteleri içindeki mevcut kısıtlama sitelerini (EcoRI, NheI, XhoI ve NcoI) ayrıntılı olarak açıklar. Yanaşma hattının yapısı da tasvir edilir. (B) Geçiş adımı, sürücü hattından erkeklerin (GAL4 sürücüsünü 3xP3 promotörü tarafından yönlendirilen bir ilgi verici ve eCFP tarafından taşıyan) ve yanıtlayıcı çizgiden kadınların (bir UAS promotörü ve 3xP3 promotörü tarafından kontrol edilen bir eYFP işaretleyicisi tarafından kontrol edilen ilgi genini veya saç tokası yapısını taşıyan) kullanımını göstermektedir. (C) GAL4’ün B’deki haçın soyuna ilgi geninin itici ifadesinin diyagramsal bir gösterimi ve değerlendirilen tipik fenotiplerden bazılarının bir listesi. Kısaltmalar: Çoklu Klonlama Bölgesi (MCS), Rekombinoz aracılı kaset değişimi (RMCE), Upstream Aktivatör Sırası (UAS), gelişmiş sarı floresan protein (eYFP), gelişmiş siyan floresan protein (eCFP). Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
Daha doğrusal yaklaşımlara göre belirgin avantajlara sahip olan GAL4-UAS sisteminin iki partili doğasını sağlayan haçların kullanılmasıdır. Örneğin, her promotör/gen kombinasyonu için yeni bir transgenik hat oluşturulması ve sürdürülmesi gerekiyorsa, sürücü ve yanıtlayıcı hatlarının birçok kombinasyonu uygulanabilir olacaktır. Daha da önemlisi, doğrusal bir sistemde oluşturulması/sürdürülmesi zor olan ifadeleri bozunduğunda ölümcül veya steril fenotip üreten genlerin analizine izin verir. Bu tür ölümcül fenotipler, gen fonksiyonuna ve mekansal ifadeye bağlı olarak tüm gelişim aşamalarında ortaya çıkabilir, ancak en sık embriyonik gelişim sırasında gözlenir. Sivrisinek embriyo gelişimini görselleştirmek, yumurtaları kaplayan opak koronun temizlenmesini gerektirir. Trpiš (1970)21 ve Kaiser ve ark. (2014)22’de açıklanan yöntemleri izleyerek, yapısal bütünlüğü korurken embriyoları düzeltmek için kullandığımız protokolleri ve mikroskobik görselleştirme ve görüntülemeye izin veren endochorion’u temizlemek için ağartmamızı açıklıyoruz.
