Before starting prepare:
- Wandering yhird instar Drosophila larvae
- HL3.1 (Modified hemolymph-like) solution
- Sylgard (transparent silicone rubber) dissection plates prepared in small (35 x 10 mm) plastic petri dishes using methods described by Brent and McCabe (2008)3.
- Cut dissection pins short
- Stimulating electrode pipettes
- Sharp recording pipettes
HL3 Solution:
- During dissections and electrophysiological experiments, larva are immersed in HL3.1 solution2 that contains (in mM): 70 NaCl, 5 KCl, 4 MgCl2, 10 NaHCO3, 5 trehalose, 115 sucrose, and 5 HEPES, pH 7.2.
- Concentrations of CaCl2 are added to the HL3.1 solution to provide the required concentration of extracellular Ca2+.
- Larval dissections are performed at low Ca2+ concentrations (to avoid muscle contraction) using HL3.1 + 0.25 mM CaCl2, kept on ice.
- Electrophysiological experiments are typically recorded in HL3.1 + 1 mM CaCl2, but this can be adjusted from 0.4 – 1 mM Ca2+ as required.
- The HL3.1 solution is filter sterilized before use, and stored at 4˚C.
Preparation of stimulating and recording pipettes:
- Recording and stimulating pipettes are pulled with a Sutter P-2000 Laser based micropipette puller using the following settings: Heat = 390, Filament = 4, Velocity = 35, Delay = 200 and Pull = 0.
- Stimulating electrodes are prepared from thin-walled borosilicate glass capillaries with wide ends that were slightly bigger than the width of the severed motor axons. The shaped ends are firepolished to give a smooth finish (using a Narashiga pipette polisher), to minimize damage to the nerve. Stimulating electrodes are filled with bath solution (HL3.1 + 1 mM Ca2+).
- The recording electrodes are prepared from 1.2mm borosilicate glass, which is pulled to form a sharp pipette (30–60 mΩ), and filled with 3M KCl.
Part 1: Dissection of Drosophila Larvae
- Third instar wandering larvae are dissected to expose the muscles in the body wall as described previously4.
- Dissections are performed in HL3.1 + 0.25 mM Ca2+ on small silicone plates (35 x 10 mm). HL3.1 solution must be ice-cold for dissections in order to anaesthetize the larva, and is kept on ice before and during dissections.
- Larvae are dissected using methods described by Brent and McCabe (2008), that is modified for electrophysiology experiments by adding 0.25 mM Ca2+ to the HL3.1 dissection solution, and using short dissection pins (~2 mm in length) that are less likely to hit the microscope objective and electrodes during an experiment.
- Following dissection, the motor axons of the larvae are severed. This is done by gently holding the CNS and raising it slightly so the peripheral nerves that innervate the muscles posterior to the ventral ganglion can be cut without damaging the muscles (Figure 1). The brain is then removed.
- The dissected larval preparation is washed twice with HL3.1 + 1 mM Ca2+.
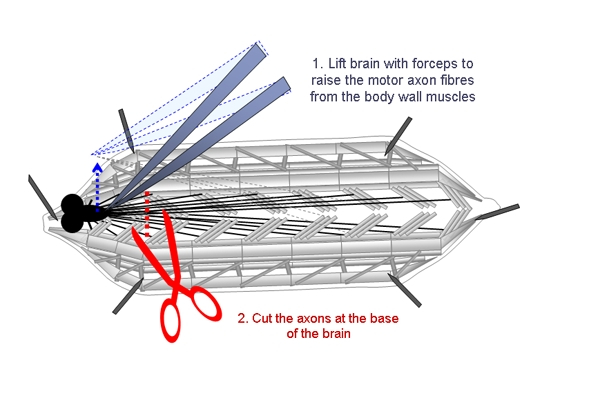
Figure 1. Schematic showing the steps involved in cutting the motor axons of dissected larvae. The CNS is lifted with forceps and the motor axons are cut at the base of the brain.
Part 2: Intracellular recordings from larval muscle cells.
- Place the dissection plate on the electrophysiology microscope and submerge the prep with HL3.1 + 1.5 mM Ca2+ and fix the bath electrode so it is in contact with the solution.
- All experiments are performed on muscle 6 within the third abdominal segment (A3). Peripheral nerves that innervate the muscles are stimulated using a suction electrode5.
- Position the stimulating electrode in the dish first to avoid vibrations when the intracellular electrode is in place. Place the stimulating electrode in close vicinity to the motor neuron innervating muscle 6 and apply gentle suction until the cut nerve is inside the glass pipette. Be careful not to stretch the nerves when suction is applied. Raise the pipette slightly so it is not touching the muscle and position it so it is not pulling on the cut nerve fibers (Figure 2).
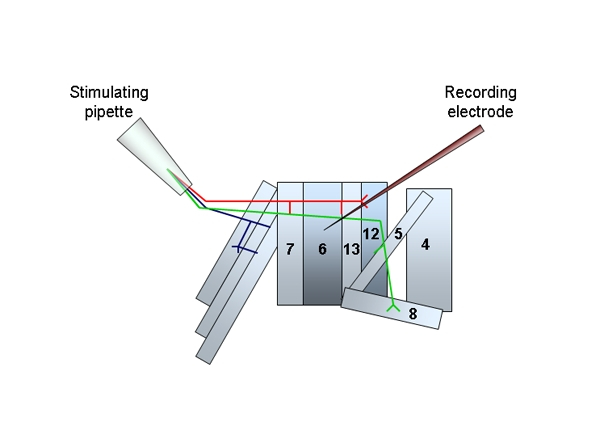
Figure 2. A schematic of the musculature and innervating motor neurons of one abdominal segment from the Drosophila larva. The position of the stimulating pipette and severed nerves and the recording electrode in position at muscle 6 are illustrated. - Intracellular recordings are made with sharp microelectrodes filled with 3 M KCl. Position the recording electrode above the centre of muscle 6 on the third abdominal segment (A3) and adjust the input offset so it reads zero for the bath solution. Slowly lower the electrode and approach the muscle under high optical magnification until it touches the muscle surface. Watch the oscilloscope to confirm that the muscle has been penetrated. The resting membrane potential should be at least -60 mV, or the animal should not be used. Sporadic miniature endplate potentials should now be visible. Leave the cell to stabilize for one minute before starting to record.
- Changes in membrane potential are detected with an Axon HS-2A head stage and an Axoclamp 2B amplifier and recorded with Clampex v 8.2.0.235 (Axon Instruments). The Axoclamp 2B is interfaced to a computer that runs pCLAMP software and works in conjunction with the Digidata 1322A interface.
- Cells were recorded for 3 minutes without any stimuli to measure mEJP responses. Traces were analysed using Mini analysis software (Synaptosoft, v 6.0.3) and the mEJP amplitude and frequency determined.
Nerve Stimulation:
- The severed end of the motor neuron is stimulated with a series of square voltage pulses (0.3 ms duration) at an intensity that is sufficient for both motor axons, to evoke consistent responses in the whole muscle.
- The stimulus is generated with the Master-8 pulse generator, which is programmed to deliver pulses continuously according to the duration and interval times.
- Allow five seconds between stimuli for synaptic recovery at the NMJ.
- When determining the strength of the stimulus, begin with the lowest level and increase the intensity slowly over several trials. The minimum stimulus intensity that results in an evoked response should be used in experiments.
- When the appropriate stimulus intensity is reached, there should be one compound EJP and one muscle contraction evoked by the stimulus.
- Record at least 10 evoked potentials from each muscle and average the amplitudes.
Part 3: Representative Results
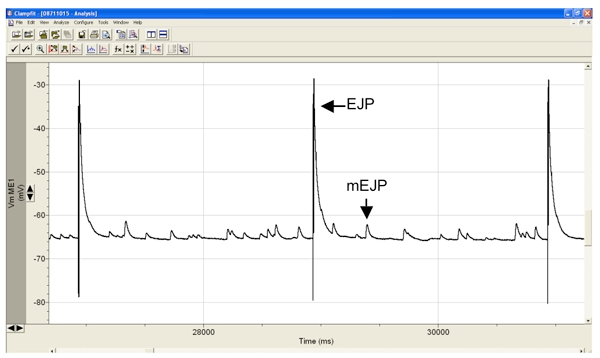
Figure 3. Representative intracellular recording from muscle 6 showing the evoked EJP’s is response to electrical stimulation of the segmental nerve, and the sporadic minature endplate potentials, or mEJP’s. EJP amplitudes in muscle 6 of healthy wild-type larvae, such as Canton S, are typically around 40 mV and the mEJP amplitudes between 1-3 mV.
