Staining for eGFP should begin showing weak signal in primary motor neurons approximately 72 hr after injecting PRV152 into muscle. The replication and transsynaptic transport of virus are titer- and time-dependent4. Approximately 90 hr after injection, eGFP staining will reveal robust signal in 2nd order infected cells. Longer survival times will reveal 3rd and higher order cells but survival times are limited by the lethality of PRV at approximately 5 days after inoculation.
Figure 1a shows neurons infected with PRV152 expressing eGFP in nucleus ambiguus, which houses the laryngeal motor neurons, 94 hr after injection of PRV152 into two of the seven laryngeal muscles. Because the virus entered the brain through neurons that innervate the infected muscles, other phonatory motor neuron pools, such as the hypoglossal nucleus, do not express eGFP (Figure 1b).
Figure 2 shows retrograde transport of the virus to 2nd order brainstem neurons subsequent to primary infection in nucleus ambiguus. The injection was unilateral, and the resulting pattern of label in the brainstem shows infection of the ipsilateral nucleus ambiguus and solitary nucleus (Figure 2a). Reticular interneurons were also infected and stained strongly for eGFP (Figure 2b). These interneurons connect various structures, including the phonatory motor neuron pools and the contralateral nucleus ambiguus; however, their efferent targets remain unlabeled because the virus only spreads in the retrograde direction.
Figure 3 shows infected premotor neurons in the motor cortex expressing eGFP contralateral to the peripheral injection site. Because PRV152 spreads through multi-synaptic circuits impinging on laryngeal motor neurons in nucleus ambiguus, it can be assumed that only the population of cells that compose the motor cortical representation of the laryngeal musculature were infected. The lack of eGFP signal in the rest of the section shown (and most of the forebrain not shown) demonstrates the specificity of the labeling.
Figure 4 shows BDA-labeled axons in the brainstem at the level of nucleus ambiguus. The axons make contact with the CTb-positive motor neurons retrogradely labeled by injections into laryngeal muscles. Axons are shown forming varicosities and putative terminal boutons near the dendrites and soma of laryngeal motor neurons. Further experiments using electron microscopy or electrophysiology are required to determine whether these varicosities represent synaptic boutons.
Figure 5a shows a bilateral injection of BDA placed into the laryngeally connected motor cortex as identified by retrograde transynaptic tracing with PRV152. Because BDA is not directionally specific, both afferent and efferent connections may be labeled. For example, both a projection field (anterograde) and cell bodies (retrograde) label are seen in the thalamus (Figure 5b).
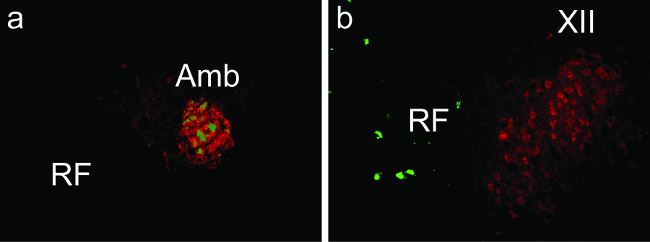
Figure 1: Neurons Infected with PRV152 Expressing eGFP (green) and Choline Acetyltransferase Immunopositive Motoneurons (red) in the (a) Ambiguus (Amb) and (b) Hypoglossal (XII) Nuclei of the Mouse Brainstem 94 hr after Injection of PRV152 into the Cricothyroid and Lateral Cricoarytenoid Laryngeal Muscles. RF = reticular formation. Please click here to view a larger version of this figure.
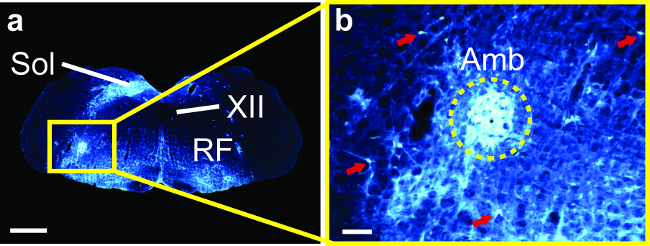
Figure 2: Psuedorabies Virus (PRV152) Infection in the Brainstem of a Mouse 86 hr after Injecting PRV152 into the Cricothyroid and Lateral Cricoarytenoid Laryngeal Muscles. (a) Neurons expressing enhanced green fluorescent protein (white; colors inverted from original brightfield images of stained sections) in nucleus ambiguus (Amb) ipsilateral to the injected muscle, the surrounding reticular formation (RF) and the solitary nucleus (Sol), but not the hypoglossal nucleus (XII). (b) Amb and reticular interneurons at higher magnification (red arrows, cell bodies in RF). Scale bars: 1 mm for a; 100 µm for b. (Modified from Arriaga et al. 2012). Please click here to view a larger version of this figure.
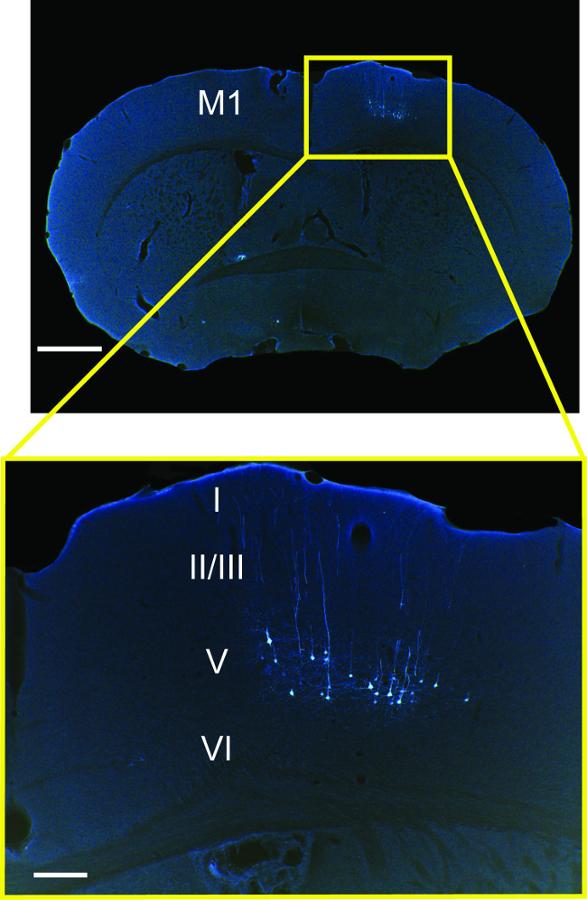
Figure 3: Cortical Pyramidal Neurons Expressing eGFP (white; colors inverted from original brightfield images of stained sections) in Cortical Layer V of M1 following Injection of Psuedorabies Virus (PRV152) into the Cricothyroid and Lateral Cricoarytenoid Laryngeal Muscles. Scale bars: 1 mm for main panel; 200 µm for inset. (Modified from Arriaga et al. 2012).
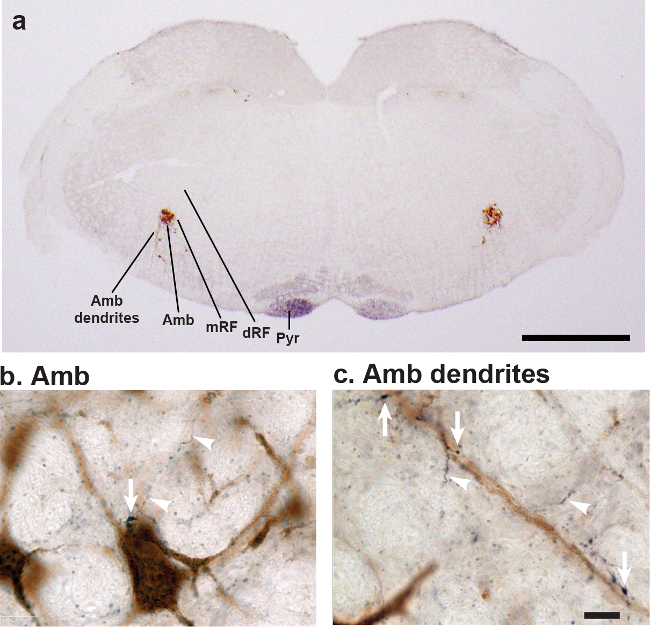
Figure 4: M1 Axons in the Brainstem. (a) Low power of a coronal brainstem section containing CTb-labeled motor neurons in Amb (brown) from an injection in laryngeal muscles and M1 axons (black) from an injection of BDA into M1. BDA labelled axons can be seen in the cortico-pyramidal (Pyr) track. Abbreviations: Amb, nucleus ambiguus; Pyr, pyramids; mRF, reticular formation directly medial to Amb; dRF, reticular formation dorsal to Amb. (b) High magnification of BDA labeled axon (black, arrow heads) from M1 that makes contact (arrow) onto a Amb motor neurons (CTb brown). (c) M1 axons (black) running along (arrows) and near (arrow heads) a large Amb motor neuron dendrite that radiates out from Amb. Please click here to view a larger version of this figure.
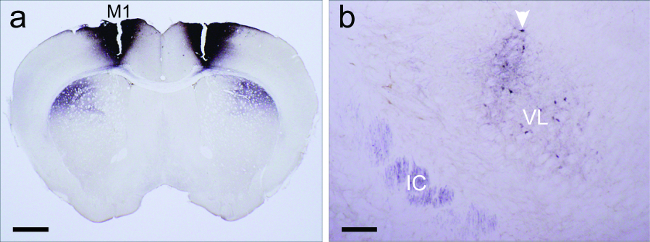
Figure 5: Injection site and additional bidirectional connections revealed by BDA injections not seen with PRV transynaptic tracing. (a) Bilateral biotinylated dextran amine (BDA) injections (black) in primary motor cortex (M1) revealed a dense terminal projection field (black) in the underlying striatum. (b) Cortical axons from M1 terminate in the ventral lateral nucleus of the thalamus (VL); A cluster of thalamic cells (dark brown and marked by white arrow head) that project back to the M1 singing region is also observed in VL. The internal capsule (IC) has labeled axons coming from the cortex. Scale bars: 1 mm for a; 200 µm for b. (Modified from Arriaga et al. 2012). Please click here to view a larger version of this figure.
