Предполагается, что геномы млекопитающих содержат около 20000 белок-кодирующих генов. Альтернативный сплайсинг и посттрансляционные модификации дальнейшего увеличения белка репертуара. Белки имеют модульную конструкцию 1 и часто содержат несколько доменов взаимодействия, которые позволяют их набор в различные белковых комплексы и их участие в многочисленных клеточных процессах 2. Одним из примеров является многофункциональный белок под названием p120ctn. p120ctn кодируется геном Ctnnd1 и состоит из большого центрального домена армадилл повтора в окружении N-концевой области и С-концевой. Домен армадилла из p120ctn связывается с высоко консервативным доменом околомембранного классических кадхеринов, которые участвуют в межклеточной адгезии, но она также связывается с репрессором транскрипции Kaiso. N-концевой домен p120ctn взаимодействует с различными киназ, фосфатаз, небольшие RhoGTPases, и ассоциированный с микротрубочками рroteins 3. Интересно отметить , что в результате альтернативного сплайсинга, p120ctn изоформы могут быть получены из четырех альтернативных стартовых кодонов 4. p120ctn изоформы 1A является самым длинным, как это переводится с большей-5' старт-кодоном и содержит полноразмерный N-концевой сегмент. В p120ctn изоформ 3 и 4, это N-концевой сегмент удаляется частично и полностью, соответственно. Понимание точной роли белков (или белкового изоформ) и их доменов в различных клеточных функциях остается проблемой.
Ген ориентации в mESCs позволяет исследовать функции белка через генетической делеции соответствующего гена и широко способствовал выявлению онтогенетически важных и болезненных значимых генов и путей. Этот прорыв в обратной генетики явилась результатом достижений в области изоляции MESC и гена ориентации за счет гомологичной рекомбинации 5 </sвверх>. Гомологичная рекомбинация представляет собой процесс, в котором происходят обмен ДНК-фрагменты между двумя аналогичными или идентичными нуклеиновыми остатками после того, как двухцепочечный (DS) разрывами ДНК. Как правило, HR является неэффективным, поскольку дц перерывы являются нечастыми. В последнее время , эффективность гомологического-направленный ген ориентации может быть увеличена с помощью сайта-специфических нуклеаз 6, 7, но , к сожалению, они склонны к вне целевых эффектов 8. Более надежный метод для того, чтобы ген таргетирование RMCE, который основан на конкретных участках систем рекомбинации, таких как Cre / LoxP или FLPe / Frt. LoxP и последовательность Frt найдены в бактериофаге Р1 и Saccharomyces CEREVISIAE, соответственно, и состоит из 34 пар оснований , в том числе последовательности асимметричных 8 пар оснований, которая определяет ориентацию сайта. С другой стороны, ориентация, например, два LoxP сайтов в участке ДНК будет определять, является ли становится вырезала floxed ДНК или яnversed при Cre-опосредованной рекомбинации 9. Кроме того, Cre может также индуцировать транслокацию, если два участка расположены на разных хромосомах. RMCE воспользовался преимуществом неспецифических сайтов рекомбинации, которые не вступают в перекрестную реакцию и внедренных в геномной локуса. В присутствии донора плазмиды, содержащая фрагмент ДНК , фланкированной один и та же неспецифических сайтами, то рекомбиназа будет вставить этот фрагмент ДНК в RMCE-совместимый геномный локус из – за двойное одновременное транслокации (рисунок 1). Здесь только правильно RMCE-ориентированные клоны могут оказать лекарственную устойчивость благодаря промотору на входящем векторе , который восстанавливает «ловушка» промотор-менее ген резистентности к неомицину (Neo R) присутствует в R26 геноме дока – клетках (фигура 1) 10, 11. Это приводит к очень высокой эффективности таргетинга, часто близкой к 100%, 11 </ SUP> 12. В заключении, на основе RMCE нацеливание имеет высокую эффективность и может быть использовано для структуры-функции исследований; Однако, это требует предварительно спроектированных геномного локуса.
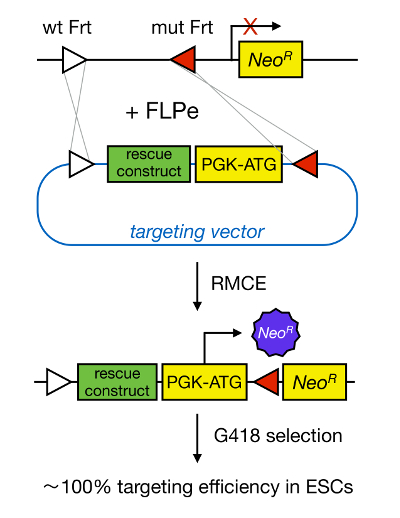
Рисунок 1. Схематическое изображение RMCE-опосредованной нацеливание. RMCE позволяет для обмена сегментов ДНК из входящего вектора направленного к определенной геномной локуса, если оба питают два неспецифических сайтов FRT (изображенные белыми и красными треугольниками). Кроме того, разработано геномный локус содержит ген без промотора и усеченной неомицин-сопротивление (Neo R). Предоставляя промотор и стартовый кодон во фрагменте ДНК входящего, только правильные события рекомбинации восстановить устойчивость к неомицину, что приводит к высокой эффективности таргетинга. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию тего фигура.
Геном инжиниринг в mESCs позволяет для генерации RMCE-совместимых мышей. В 1981 году две группы удалось захватить плюрипотентных клеток из внутренней клеточной массы (ICM) бластоцист и в поддержании их в культуре 13, 14. mESCs способны к самообновлению и дифференцировке во все типы эмбриональных и взрослых клеток, в том числе зародышевых клеток линии. Таким образом, ген ориентации в mESCs дает обратное генетические исследования путем разработки конститутивного или условного (с использованием системы / LoxP Cre) мышей KO. Тем не менее, классический способ выделения клеток мыши ES очень неэффективно. Несколько основных улучшений значительно увеличили вероятность успеха для получения линий MESC, в том числе использование определенного сывороточного замещения (SR) среды 15, чередуя MESC среде , содержащей SR и фетальной бычьей сыворотки (FBS) 16, а также использование фармакоЛогические соединения , такие как pluripotin или 2i 17. Pluripotin, небольшая синтетическая молекула, позволяет для распространения mESCs в недифференцированном состоянии в отсутствии фактора ингибировани лейкоза (LIF) и эмбриональных фибробласты мышей (MEFs) 18. Наконец, было показано , что mESCs может быть выделен с очень высокой эффективностью (близкой к 100%) , когда / ФБС протокола SR среднего чередование сочетается с LIF и pluripotin 19, 20. Эти протоколы позволяют эффективно изолировать RMCE-совместимый KO mESCs, которые впоследствии могут быть использованы для исследований структурно-функциональных.
В этой статье описан способ, который дает возможность определить ключевые домены или остатки в пределах белка, которые отвечают за специфических клеточных процессов. С этой целью трубопровод передовых технологий, позволяющих эффективно изоляции мЭСК, ориентации вектора сборки и мЭСК нацеливания было создатьд. Как таковые, большие панели с изоформы белка, мутанты доменов и вниз по течению эффекторов могут быть введены в KO mESCs и могут быть оценены по их способности , чтобы спасти в пробирке KO фенотип.
