הערכה היא כי הגנום יונק מכיל כ 20,000 גנים המקודדים לחלבונים. שחבור אלטרנטיבי ושינויי posttranslational להגביר עוד יותר את רפרטואר החלבון. יש חלבוני מבנה מודולרי 1 ולעתים קרובות מכילים תחומי אינטראקציה מרובים, המאפשרים הגיוס שלהם לתוך מתחמי חלבון שונים השתתפותם תהליכים תאיים רבים 2. דוגמה אחת היא חלבון רב תפקודי הנקרא p120ctn. p120ctn מקודד על ידי הגן Ctnnd1 ו מורכב מתחום חוזרים ארמדיל מרכזי גדול ולצידו ידי N-מסוף אזור הטרמינל-C. התחום ארמדיל של p120ctn נקשר לתחום juxtamembrane השמור ביותר של cadherins הקלסית, אשר מעורבים תאי תאי הידבקות, אבל זה גם נקשר מדכא תעתיק Kaiso. התחום מסוף-N של p120ctn אינטראקציה עם קינאזות שונות, phosphatases, RhoGTPases קטן, ו- p-מזוהה microtubuleroteins 3. מעניין, כתוצאה של שחבור חלופי, isoforms p120ctn ניתן להפיק ארבעה קודונים התחלה חלופה 4. 1A איזופורם p120ctn הוא הארוך ביותר, כפי שהוא מתורגם מן-5 ביותר להתחיל קודון ומכיל קטע באורך מלא N-terminal. בשנת p120ctn isoforms 3 ו 4, זה קטע N-terminal נמחק חלקית לחלוטין, בהתאמה. הבנת התפקיד המדויק של חלבונים (או isoforms חלבון) ועל התחומים שלהם פונקציות סלולריות שונות עדיין מהווה אתגר.
ג'ין מיקוד mESCs מאפשר הלימוד של הפונקציה של חלבון דרך המחיקה הגנטית של הגן המתאים ואת תרם רב לזיהוי גנים ושבילים התפתחותית חשובים ומחל-רלוונטיות. זו פריצת דרך גנטיקה הפוכה הייתה התוצאה של התקדמות בתחומי בידוד המסקלין ואת גן המיקוד בשל הומולוגי 5 </sעד>. הומולוגיים הוא תהליך שבו שברי DNA הם החליפו בין שני moieties גרעין דומה או זהה לאחר הפסקות פעמיים גדילי דנ"א (DS). בדרך כלל, HR אינו יעיל משום הפסקות dsDNA הן נדירות. לאחרונה, את היעילות של גנים בבימויו הומולוגית מיקוד יכולה להיות מוגברת באמצעות nucleases ספציפי באתר 6, 7, אבל לצערי, אלה נוטים השפעות חוץ-יעד 8. טכניקה אמינה יותר כדי לאפשר גן המיקוד הוא RMCE, אשר מבוססת על מערכות רקומבינציה ספציפי באתר כגון Cre / loxP או FLPe / FRT. LoxP ורצף FRT נמצאים P1 בקטריופאג ו שמר האפייה, בהתאמה, וכוללת 34 נ"ב, כולל רצף BP אסימטרי 8 הקובע את הכיוון של האתר. מצד השני, את הכיוון של, למשל, שני אתרי loxP בתוך מתיחת DNA יקבעו אם הדנ"א floxed הופך ניכר או inversed על רקומבינציה 9 בתיווך Cre. יתר על כן, Cre יכול גם לגרום טרנסלוקציה אם שני האתרים נמצאים על כרומוזומים שונים. RMCE מנצל אתרי רקומבינציה heterospecific כי לא צולב להגיב וכי מוטבעי מוקד הגנומי. בנוכחות פלסמיד התורם המכיל קטע DNA מוקף באותם אתרים heterospecific, את recombinase יוסיף קטע DNA הזה לתוך לוקוס גנומית תואם RMCE בגלל טרנסלוקציה פעמיים סימולטני (איור 1). הנה, רק בצורה הנכונה RMCE במיקוד שיבוטים יכולים לעבד בזכות עמידות לתרופות אמרגן על הווקטור נכנס כי משחזר "לכודים" גני אמרגן-פחות התנגדות neomycin (ניאו R) הנוכחים בגנום R26 של תאי העגינה (איור 1) 10, 11. התוצאה היא יעילות מיקוד גבוהה מאוד, לעתים קרובות קרובה 100% 11, </ sup> 12. לסיכום, מיקוד מבוסס RMCE הוא יעיל ביותר וניתן להשתמש בו ללימודים-פונקציות מבנה; עם זאת, זה דורש מוקד הגנומי טרום מהונדס.
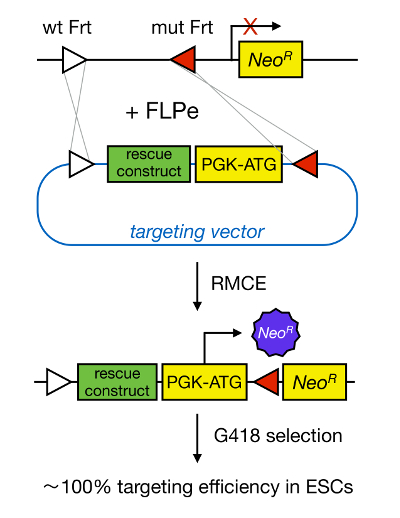
איור 1. ייצוג סכמטי של מיקוד בתיווך RMCE. RMCE מאפשר החלפת מקטעי דנ"א מתוך וקטור מיקוד הנכנסת מוקד הגנומי מוגדר אם הן נמל שני אתרי FRT heterospecific (מתואר על ידי משולשים לבנים ואדומים). בנוסף, לוקוס גנומי מהונדסים מכיל promoterless ו-התנגדות neomycin קטועה גן (ניאו R). על ידי מתן מקדם ולהתחיל קודון של קטע DNA נכנס, רק אירועי רקומבינציה נכונים לשחזר התנגדות neomycin, וכתוצאה מכך יעיל מיקוד גבוה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של tהדמות שלו.
הנדסת הגנום ב mESCs מאפשרת לדור של עכברים תואמים RMCE. בשנת 1981, שתי הקבוצות הצליחו ללכוד תאים פלוריפוטנטיים ממסת התאים הפנימית (ICM) של blastocysts ובשמירה עליהם בתרבות 13, 14. mESCs מסוגל התחדשות עצמית והבחן לתוך כל סוגי תאים עובריים מבוגרים, כולל שושלת התא הניבט. לכן, גן מיקוד mESCs מאפשר לימודים הפוכים-גנטיים באמצעות הפיתוח של מכונן או מותנים (באמצעות מערכת Cre / LoxP) עכברי KO. עם זאת, הדרך הקלאסית לבודד תאי עכבר ES הוא מאוד לא יעיל. שיפורים מרכזיים הגדילו את שיעור ההצלחה מאוד עבור קווי המסקלין נובעים, כוללים השימוש בסרום-החלפה מוגדרת (SR) בינונית 15, לסירוגין בין מדיום המסקלין המכיל סרום שור SR ו עוברי (FBS) 16, ושימוש pharmacoתרכובות לוגיות כגון pluripotin או 2i 17. Pluripotin, מולקולה סינתטית קטנה, מאפשר התפשטות של mESCs במצב מובחן בהעדר גורם מעכבות לוקמיה (LIF) ו פיברובלסטים עובריים בעכבר (MEFs) 18. לבסוף, הוכח כי ניתן לבודד mESCs עם יעילות גבוהה מאוד (קרוב ל 100%), כאשר פרוטוקול ההתחלפות בינוני SR / FBS משולב עם LIF ו pluripotin 19, 20. פרוטוקולים אלה מאפשרים בידוד יעיל של mESCs RMCE תואם KO שיכולים לשמש בהמשך למחקרים מבנה-תפקוד.
מאמר זה מתאר שיטה המאפשרת אחד כדי לזהות את התחומים או שאריות המפתח בתוך חלבון כי הם אחראים על תהליכים תאיים ספציפיים. לשם כך, צנרת של טכנולוגיות מתקדמות המאפשרות בידוד המסקלין יעיל, הרכבת וקטור מיקוד, מיקוד המסקלין הייתה ליצורד. כמו לוחות כאלה, גדולים עם isoforms חלבון, מוטציות תחום ומפעילים במורד הזרם יכול להיות הציג mESCs KO וניתן להעריך את יכולתם להציל את הפנוטיפ KO חוץ גופית.
