On estime que les génomes de mammifères contiennent environ 20 000 gènes codant pour des protéines. L'épissage alternatif et modifications post-traductionnelles augmenter encore le répertoire de protéines. Les protéines ont une structure modulaire 1 et contiennent souvent de multiples domaines d'interaction, qui permettent leur recrutement dans différents complexes de protéines et leur participation à de multiples processus cellulaires 2. Un exemple est la protéine multifonctionnelle appelée p120ctn. p120ctn est codée par le gène Ctnnd1 et se compose d'un grand domaine de répétition de tatou centrale flanquée par une extrémité N-terminale et une région C-terminale. Le domaine de tatou p120ctn se lie à un domaine juxtamembranaire hautement conservée de cadhérines classiques, qui sont impliqués dans l'adhésion cellulaire des cellules, mais il se lie également au répresseur de la transcription Kaiso. Le domaine N-terminal de p120ctn interagit avec différentes kinases, les phosphatases, les petites GTPases Rho et associée aux microtubules proteins 3. Il est intéressant, en raison de l' épissage alternatif, isoformes p120ctn peuvent être générés à partir de quatre codons de départ alternatifs 4. p120ctn isoforme 1A est la plus longue, elle est traduite à partir de la plus 5' codon de départ et contient la longueur complète segment N-terminal. Dans p120ctn isoformes 3 et 4, ce segment N-terminal est supprimé partiellement et complètement, respectivement. Comprendre le rôle précis des protéines (ou isoformes de protéines) et leurs domaines dans différentes fonctions cellulaires reste un défi.
Le ciblage de gènes dans mESCs permet l'étude de la fonction d'une protéine à travers la deletion génétique du gène correspondant et a largement contribué à l'identification des gènes et les voies de développement importants et importantes maladies. Cette percée en génétique inverse a été le résultat des progrès dans les domaines de l' isolement MESC et ciblage de gène en raison de la recombinaison homologue 5 </sup>. La recombinaison homologue est un procédé dans lequel des fragments d'ADN sont échangés entre deux fragments nucléiques similaires ou identiques après cassures de l'ADN (DS) double brin. En règle générale, HR est inefficace parce que les pauses dsDNA sont rares. Récemment, l'efficacité de gène dirigé vers une homologie de ciblage pourrait être augmentée en utilisant des nucléases spécifiques à un site 6, 7, mais , malheureusement, elles sont sujettes à des effets hors-cible 8. Une technique plus fiable pour permettre le ciblage de gène est CREM, qui est basée sur des systèmes de recombinaison spécifiques au site tels que Cre / loxP ou FLPE / FRT. LoxP et la séquence Frt se trouvent dans le bactériophage P1 et Saccharomyces cerevisiae, respectivement, et se composent de 34 pb, comprenant une séquence de 8 pb asymétrique qui détermine l'orientation du site. D'autre part, l'orientation, par exemple, deux sites loxP dans un tronçon d'ADN déterminera si l'ADN floxés devient excisé ou inversed lors d'une recombinaison à médiation par Cre 9. En outre, la Cre peut également induire une translocation si deux sites sont situés sur des chromosomes différents. CREM profite des sites de recombinaison hétérospécifiques qui ne sont pas provoquer une réaction croisée et qui sont incorporés dans un locus génomique. En présence d'un plasmide donneur qui contient un fragment d'ADN flanqué par les mêmes sites hétérospécifiques, la recombinase va insérer ce fragment d'ADN dans le locus génomique compatible CREM en raison de translocation à double simultanée (Figure 1). Ici, seuls correctement clones CREM ciblés peuvent rendre la résistance aux médicaments grâce à un promoteur sur le vecteur entrant qui restaure un « piégé », sans promoteur gène de résistance à la néomycine (Neo R) présente dans le génome R26 des cellules d'accueil (Figure 1) 10, 11. Il en résulte une efficacité de ciblage très élevé, souvent près de 100% 11, </ sup> 12. En conclusion, le ciblage par CREM-est très efficace et peut être utilisé pour les études de structure-fonctions; cependant, il a besoin d'un locus génomique précalculé.
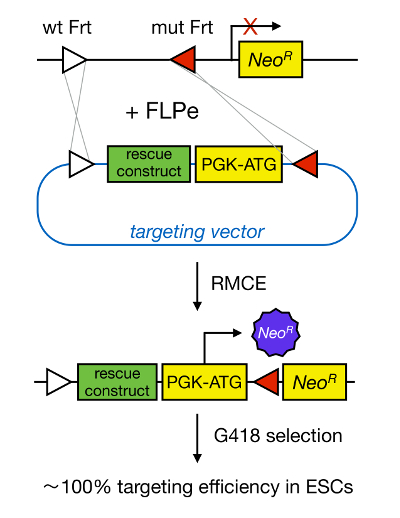
Figure 1. Représentation schématique de ciblage à médiation par CREM. CREM permet l'échange de segments d'ADN à partir d'un vecteur de ciblage entrant à un locus génomique définie si les deux hébergent deux sites hétérospécifiques Frt (représentés par des triangles blancs et rouges). En outre, le locus génomique conçu contient un promoteur et un gène de résistance à la néomycine tronquée (Neo R). En fournissant un promoteur et codon de départ dans le fragment d'ADN entrant, seuls les événements de recombinaison restauration correcte résistance à la néomycine, entraînant des rendements élevés de ciblage. S'il vous plaît cliquer ici pour voir une version plus grande de tsa figure.
L'ingénierie des génomes en mESCs permet la génération de souris compatible CREM. En 1981, deux groupes ont réussi à capturer des cellules pluripotentes à partir de la masse cellulaire interne (ICM) de blastocystes et à les maintenir en culture 13, 14. mESCs sont capables d'auto-renouvellement et de différenciation dans tous les types de cellules embryonnaires et adultes, y compris la lignée germinale. Par conséquent, le ciblage génique dans mESCs permet des études génétiques inverse par le développement de constitutifs ou conditionnels (en utilisant le système Cre / LoxP) souris KO. Cependant, la manière classique pour isoler des cellules ES de souris est très inefficace. Plusieurs améliorations importantes ont fortement augmenté le taux de succès pour dériver des lignes Mesc, y compris l'utilisation d'un sérum de remplacement défini (SR) moyen 15, en alternant entre milieu MESC contenant du sérum bovin SR et fœtal (FBS) 16, et l'utilisation de pharmacocomposés logiques tels que pluripotin ou 17 2i. Pluripotin, une petite molécule synthétique, permet la propagation de mESCs dans un état indifférencié en l'absence de facteur inhibiteur de leucémie (LIF) et les fibroblastes embryonnaires de souris (MEF) 18. Enfin, il a été démontré que mESCs peuvent être isolés avec un rendement très élevé (proche de 100%) lorsqu'une SR / FBS protocole moyen d'alternance est combiné avec du LIF et pluripotin 19, 20. Ces protocoles permettent l'isolement efficace de KO mESCs de CREM compatibles qui peuvent ensuite être utilisés pour des études structure-fonction.
Le présent document décrit une méthode que l'on permet d'identifier les domaines clés ou les résidus dans une protéine qui sont responsables des processus cellulaires spécifiques. À cette fin, un pipeline de technologies de pointe qui permettent l'isolement MESC efficace, le ciblage ensemble vecteur, et le ciblage MESC a été créeré. En tant que tel, de grands panneaux avec des isoformes de protéines, des mutants de domaine et effecteurs en aval peuvent être introduits dans KO mESCs et peuvent être évalués pour leur capacité à sauver le phénotype KO in vitro.
