Geschat wordt dat zoogdieren genomen bevatten ongeveer 20.000 eiwitcoderende genen. Alternatief splitsen en posttranslationele modificaties verder te verhogen het eiwit repertoire. Eiwitten een modulaire constructie 1 en bevatten vaak meerdere interactie-domeinen, waardoor hun rekrutering in verschillende eiwitcomplexen en hun deelname aan verschillende celprocessen 2 mogelijk. Een voorbeeld is de multifunctionele eiwit genaamd p120ctn. p120ctn wordt gecodeerd door het gen Ctnnd1 en bestaat uit een groot centraal armadillo herhalende domein geflankeerd door een N-terminale en C-terminale gebied. De armadillo domein van p120ctn bindt aan een sterk geconserveerd juxtamembraandomein klassieke cadherinen, die betrokken zijn bij cel-cel adhesie, maar bindt ook aan de transcriptionele repressor Kaiso. Het N-terminale domein van p120ctn interageert met andere kinasen, fosfatasen, kleine RhoGTPases en microtubule geassocieerde pProteins 3. Interessant is dat als gevolg van alternatieve splicing, p120ctn isovormen kunnen worden gegenereerd uit vier alternatieve startcodons 4. p120ctn isovorm 1A is het langst, zoals vertaald vanaf de meest 5' startcodon en bevat de volledige lengte N-eindstandige segment. In p120ctn isovormen 3 en 4, wordt dit N-eindstandige segment gedeeltelijk en volledig respectievelijk verwijderd. Inzicht in de precieze rol van eiwitten (of eiwit isovormen) en hun domeinen in verschillende cellulaire functies blijft een uitdaging.
Gen-targeting in mESCs maakt het bestuderen van de functie van een proteïne door genetische deletie van het overeenkomstige gen en heeft een grote bijdrage tot de identificatie van ontwikkelings belangrijk en ziekte-relevante genen en pathways. Deze doorbraak in reverse genetics was het gevolg van de ontwikkelingen op het gebied van mES isolatie en gene targeting als gevolg van homologe recombinatie 5 </sup>. Homologe recombinatie is een werkwijze waarbij DNA-fragmenten worden uitgewisseld tussen twee gelijkaardige of identieke nucle- resten na dubbelstrengs (ds) DNA-breuken. Normaal gesproken, HR is inefficiënt omdat dsDNA pauzes zijn zeldzaam. Onlangs zou de efficiëntie van homologie gerichte gerichte mutagenese worden verhoogd middels plaatsspecifieke nucleasen 6, 7, maar helaas zijn deze gevoelig voor off-doeleffecten 8. Een meer betrouwbare techniek voor gen targeting inschakelen is RMCE, die gebaseerd is op plaatsspecifieke recombinatie systeem zoals Cre / loxP of FLPe / Frt. LoxP en Frt sequentie in bacteriofaag P1 en Saccharomyces cerevisiae, respectievelijk, en bestaan uit 34 bp, waaronder een asymmetrische 8 bp sequentie die de oriëntatie van de site bepaalt. Anderzijds de oriëntatie van, bijvoorbeeld twee loxP plaatsen in de DNA strook bepaalt of de floxed DNA wordt uitgesneden of inversed upon Cre gemedieerde recombinatie 9. Bovendien kan Cre ook translocatie induceert wanneer twee plaatsen zich op verschillende chromosomen. RMCE maakt hiervan heterospecifiek recombinatie locaties die niet kruisreageren en die zijn ingebed in een genomische locus. In aanwezigheid van een donor plasmide dat een DNA-fragment geflankeerd door dezelfde heterospecifiek plaatsen bevat, zal de recombinase dit DNA-fragment in te voegen in de RMCE-compatibele genomische locus verdubbelen als gevolg van gelijktijdige translocatie (figuur 1). Hier kunnen alleen correct RMCE gerichte klonen render geneesmiddelresistentie door een promotor van de inkomende vector die herstelt een "gevangen" promoter-less neomycine resistentiegen (neo R) in de R26 genoom van het docking cellen (figuur 1) 10, 11. Dit resulteert in een zeer hoge targeting efficiency, vaak in de buurt van 100% 11, </ sup> 12. Concluderend-RMCE gebaseerde targeting is zeer efficiënt en kan worden gebruikt voor structuur-functie studies; echter, het vereist een prefab genomische locus.
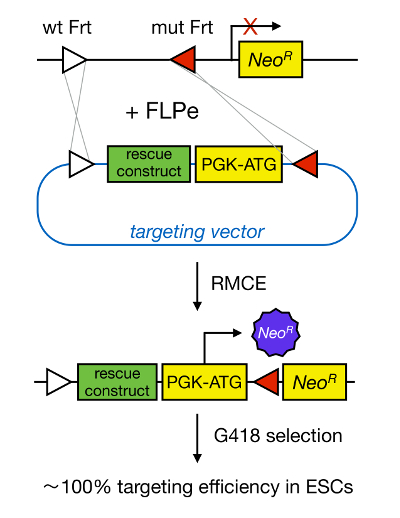
Figuur 1. Schematische weergave van RMCE-gemedieerde targeting. RMCE maakt de uitwisseling van DNA-segmenten uit een inkomend richtende vector op een bepaalde genomische locus als zowel haven heterospecifiek twee FRT-plaatsen (aangegeven door witte en rode driehoekjes). Bovendien, de gemanipuleerde genome locus bevat een promoter en afgeknot neomycine-resistentie (Neo R) gen. Door een promoter en startcodon in het inkomende DNA-fragment, enige juiste recombinatiegebeurtenissen herstellen neomycine resistentie, wat resulteert in hoge rendementen targeting. Klik hier om een grotere versie van t bekijkenzijn figuur.
Genome Engineering in mESCs maakt het mogelijk om het genereren van RMCE-compatibele muizen. In 1981, twee slaagden in het vastleggen van pluripotente cellen van de binnenste celmassa (ICM) van blastocysten en het handhaven in kweek 13, 14. mESCs in staat zijn zelfvernieuwing en differentiatie tot alle embryonale en volwassen cellen, inclusief de geslachtscellen-cellijn. Daarom gen-targeting in mESCs maakt reverse genetische studies door de ontwikkeling van constitutieve of voorwaardelijke (via het Cre / loxP systeem) KO muizen. Echter, de klassieke manier om de muis ES-cellen te isoleren is zeer inefficiënt. Aantal belangrijke verbeteringen sterk verhoogd succes voor het afleiden mES lijnen, inclusief het gebruik van een bepaald serum-vervanging (SR) -medium 15, afwisselend mES medium met SR en foetaal runderserum (FBS) 16 en het gebruik farmacologische verbindingen zoals pluripotin of 17 2i. Pluripotin, een klein synthetisch molecuul, maakt de voortplanting van mESCs in een ongedifferentieerde toestand bij afwezigheid van leukemie remmende factor (LIF) en muis embryonale fibroblasten (MEF) 18. Tenslotte is aangetoond dat mESCs kan worden geïsoleerd met een zeer hoog rendement (bijna 100%) wanneer een SR / FBS medium afwisseling protocol wordt gecombineerd met LIF en pluripotin 19, 20. Deze protocollen maken de efficiënte isolatie van RMCE-compatibele KO mESCs die vervolgens kunnen worden gebruikt voor structuur-functie studies.
Dit artikel beschrijft een werkwijze die het mogelijk maakt om de belangrijkste domeinen of resten te identificeren in een eiwit dat verantwoordelijk is voor specifieke cellulaire processen. Te dien einde, een pijpleiding van geavanceerde technologieën die het mogelijk maken een efficiënte mES isolatie, targeting vector assemblage en mES targeting was creërend. Als zodanig grote panelen met eiwit isovormen, domain mutanten en downstream effectoren in KO mESCs kan worden ingevoerd en kan worden geëvalueerd op hun vermogen om de in vitro KO fenotype te redden.
