Es wird geschätzt, dass etwa 20.000 Säugergenomen Protein-kodierenden Gene enthalten. Alternatives Spleißen und posttranslationale Modifikationen erhöhen die Proteinrepertoires. Proteine haben einen modularen Aufbau 1 und häufig mehrere Wechselwirkungsdomänen enthalten, die ihre Einstellung 2 in verschiedene Proteinkomplexe und ihre Beteiligung an mehreren zellulären Prozesse ermöglichen. Ein Beispiel dafür ist das multifunktionale Protein namens p120ctn. p120ctn wird durch das CTNND1 Gen codiert und besteht aus einer großen zentralen Domäne Armadillo – Repeat flankiert von einem N-terminalen und C-terminalen Region. Die Armadillo-Domäne von p120ctn bindet an eine hoch konservierten Juxtamembrandomäne der klassischen Cadherine, die in Zell-Zell-Adhäsion beteiligt sind, aber es bindet auch an den Repressor Kaiso. Die N-terminale Domäne von p120ctn interagiert mit verschiedenen Kinasen, Phosphatasen, kleinen RhoGTPasen und Mikrotubuli-assoziierte proteins 3. Interessanterweise als Ergebnis des alternativen Spleißens, p120ctn Isoformen können 4 aus vier alternativen Startcodons erzeugt werden. p120ctn Isoform 1A ist die längste, wie es aus dem am meisten 5' translatiert wird Startcodon und enthält die Volllängen-N-terminales Segment. In p120ctn Isoformen 3 und 4 ist dieses N-terminale Segment teilweise gelöscht und vollständig sind. die genaue Rolle von Proteinen (oder Protein-Isoformen) und die Domänen in verschiedenen zellulären Funktionen zu verstehen, bleibt eine Herausforderung.
Gen-Targeting in mESCs ermöglicht die Untersuchung der Funktion eines Proteins durch die genetische Deletion des entsprechenden Gens und hat weitgehend dazu beigetragen, die Identifizierung von entwicklungs wichtig und krankheitsrelevanten Genen und Wegen. Dieser Durchbruch in der reversen Genetik war das Ergebnis der Fortschritte in den Bereichen der MESC Isolierung und Gene aufgrund homologer Rekombination Targeting 5 </sup>. Homologe Rekombination ist ein Verfahren, bei der DNA-Fragmenten zwischen zwei ähnlichen oder identischen Nukleins Einheiten nach doppelsträngige (ds) DNA-Brüchen ausgetauscht werden. Normalerweise ist HR ineffizient, weil dsDNA Pausen selten sind. Seit kurzem wird die Effizienz der Homologie-directed gene targeting könnte unter Verwendung von ortsspezifischen Nucleasen erhöht 6, 7, aber unglücklicherweise sind diese anfällig für Nebeneffekte 8. Eine zuverlässigere Technik zu ermöglichen, Gen-Targeting ist RMCE, die auf ortsspezifische Rekombinationssysteme, wie Cre / loxP oder FLPe / FRT-basiert. LoxP und Frt – Sequenz werden in Bakteriophage P1 und Saccharomyces cerevisiae, bzw., und aus 34 bp, einschließlich einer asymmetrischen 8 – bp – Sequenz gefunden, die die Orientierung der Website festlegt. Auf der anderen Seite ist die Orientierung von, zum Beispiel, zwei loxP-Stellen innerhalb einer DNA-Strecke werden bestimmen, ob die floxed DNA exzidiert wird oder i9 nversed nach Cre-vermittelter Rekombination. Darüber hinaus kann Cre induziert auch eine Translokation, wenn zwei Standorte auf verschiedene Chromosomen befinden. RMCE nutzt artfremden Rekombinationsstellen, die nicht kreuzreagieren und das in einer genomischen Locus eingebettet sind. In Anwesenheit eines Donator – Plasmid , das ein DNA – Fragment mit den gleichen hetero Stellen flankierte enthält, fügt die Rekombinase dieses DNA – Fragment in den kompatibelen RMCE genomischen Locus durch doppelte versetzte Translokation (Abbildung 1). Hier wird nur richtig RMCE gezielte Klone können drug resistance dank eines Promotors auf dem ankommenden Vektor rendern , die ein „gefangen“ , stellt promotorlose Neomycinresistenzgen (Neo R) in der R26 Genom der Docking – Zellen (Abbildung 1) 10, 11. Daraus ergibt sich eine sehr hohe Targeting Effizienz, oft nahe 100% 11 </ sup> 12. Abschließend ist RMCE basierte Targeting sehr effizient und kann für Struktur-Funktionen Studien verwendet werden; Jedoch erfordert es eine vorgefertigte genomischen Locus.
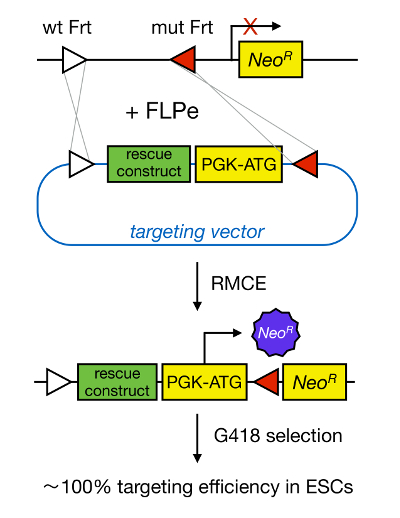
Abbildung 1 : Schematische Darstellung der RMCE Vermittelte Targeting. RMCE ermöglicht den Austausch von DNA-Segmenten aus einem ankommenden Targeting-Vektor zu einem definierten genomischen Locus, wenn sowohl zwei hetero FRT-Stellen Harbor (dargestellt durch weiße und rote Dreiecke). Darüber hinaus enthält das manipulierte genomischen Locus und ein promotorloses Neomycin-Resistenz verkürztes (Neo R) -Gen. Durch die Bereitstellung von Wiederherstellung einen Promotor und Startcodon in dem ankommenden DNA-Fragmente, nur korrekte Rekombinationsereignisse Neomycin-Resistenz, was zu hohen Effizienzen Targeting. Bitte klicken Sie hier , um eine größere Version t anzuzeigenseine Figur.
Genomtechnik in mESCs ermöglicht die Erzeugung von RMCE-kompatible Mäuse. Im Jahr 1981 gelang es zwei Gruppen von pluripotenten Zellen der inneren Zellmasse (ICM) von Blastozysten in Erfassung und in Kultur 13, 14 aufrechtzuerhalten. mESCs der Lage ist, die Selbsterneuerung und Differenzierung in alle Arten von embryonalen und adulten Zellen, einschließlich der Keimzelllinie. Daher Gene in mESCs Targeting ermöglichen Reverse genetische Studien über die Entwicklung von konstitutiven oder bedingten KO-Mäusen (die Cre / loxP-System verwendet wird). Allerdings ist die klassische Art und Weise Maus-ES-Zellen zu isolieren, ist sehr ineffizient. Verschiedene Verbesserungen haben große stark erhöht die Erfolgsrate für die Ableitung MESC Linien, einschließlich der Verwendung eines definierten serum-Ersatz (SR) Medium 15, im Wechsel zwischen MESC Medium SR und fötales Rinderserum (FBS) , 16, und die Verwendung von pharmakodynamischen enthaltendlogische Verbindungen wie pluripotin oder 2i 17. Pluripotin, ein kleines synthetisches Molekül, ermöglicht die Ausbreitung von mESCs in einem undifferenzierten Zustand in Abwesenheit von Leukämie – Hemmfaktor (LIF) und embryonalen Mausfibroblasten (MEFs) 18. Schließlich hat es sich gezeigt , dass mESCs kann mit einem sehr hohen Wirkungsgrad ( in der Nähe von 100%) isoliert werden , wenn ein SR / FBS – Medium Wechsel – Protokoll mit LIF kombiniert und pluripotin 19, 20. Diese Protokolle ermöglichen die effiziente Isolierung von RMCE-kompatibel KO mESCs, die anschließend für Struktur-Funktions-Studien verwendet werden können.
Dieses Dokument beschreibt ein Verfahren, das man die Schlüssel-Domänen oder Reste innerhalb eines Proteins zu identifizieren, ermöglicht es, die für spezifische zelluläre Prozesse verantwortlich sind. Zu diesem Zweck wird eine Pipeline von fortschrittlichen Technologien, die eine effiziente MESC Isolierung ermöglichen, Targeting-Vektor Montage und MESC Targeting war erstellend. Als solche großen Platten mit Proteinisoformen, Domain – Mutanten und Effektoren können in KO mESCs eingeführt werden können und für ihre Fähigkeit , die in – vitro – KO – Phänotyp zu retten ausgewertet werden.
