Det anslås, at pattedyrgenomer indeholder omkring 20.000 proteinkodende gener. Alternativ splejsning og posttranslationelle modifikationer yderligere øge protein repertoire. Proteiner har en modulær struktur 1 og ofte indeholde multiple interaktionsdomæner, som tillader deres rekruttering til forskellige protein-komplekser og deres deltagelse i flere cellulære processer 2. Et eksempel er det multifunktionelle protein kaldet p120ctn. p120ctn kodes af Ctnnd1 genet og består af et stort centralt bæltedyr gentagelsesdomæne flankeret af en N-terminal og en C-terminal region. Bæltedyret domæne p120ctn binder til en særdeles konserveret juxtamembrandomæne af klassiske cadheriner, som er involveret i celle-celle-adhæsion, men det binder også til den transskriptionsrepressor Kaiso. Det N-terminale domæne af p120ctn interagerer med forskellige kinaser, phosphataser, små RhoGTPases og mikrotubulus-associeret proteins 3. Interessant, som et resultat af alternativ splejsning, p120ctn isoformer kan genereres fra fire alternative startkodoner 4. p120ctn isoform 1A er den længste, da det er oversat fra den mest-5' startkodon og indeholder fuldlængde N-terminale segment. I p120ctn isoformerne 3 og 4 er dette N-terminale segment delvist og fuldstændigt henholdsvis slettet. Forstå den præcise rolle proteiner (eller protein-isoformer) og deres domæner i forskellige cellulære funktioner fortsat en udfordring.
Genmålretning i mESCs muliggør studiet af funktionen af et protein gennem genetisk deletion af det tilsvarende gen og har bredt bidraget til identifikation af udviklingsmæssigt vigtige og sygdomsrelaterede relevante gener og veje. Dette gennembrud i revers genetik var resultatet af fremskridt inden for Mesc isolation og genmålretning grund homolog rekombination 5 </sop>. Homolog rekombination er en proces, hvor DNA-fragmenter udveksles mellem to lignende eller identiske nucleinsyre-dele efter dobbeltstrenget (ds) DNA-brud. Normalt HR er ineffektivt, fordi dsDNA pauser er sjældne. Nylig kunne effektiviteten af homologi-dirigeret genmålretning øges ved anvendelse af stedspecifikke nukleaser 6, 7, men desværre disse er tilbøjelige til off-target effekter 8. En mere pålidelig teknik til at muliggøre genmålretning er RMCE, som er baseret på stedspecifikke rekombinationssteder systemer såsom Cre / loxP eller FLPe / Frt. LoxP og Frt sekvens findes i bakteriofag P1 og Saccharomyces cerevisiae henholdsvis og består af 34 bp, herunder en asymmetrisk 8 bp sekvens, der bestemmer orienteringen af sitet. På den anden side, orienteringen af for eksempel to loxP-sites inden for en DNA-strækning vil afgøre, om floxed DNA bliver udskåret eller inversed upon Cre-medieret rekombination 9. Desuden kan Cre også inducere en translokation hvis to steder er placeret på forskellige kromosomer. RMCE drager fordel af Heterospecifikke rekombinationssteder, der ikke krydsreagerer, og som er indlejret i et genomisk locus. I nærvær af et donorplasmid, der indeholder et DNA-fragment flankeret af de samme Heterospecifikke sites, vil rekombinasen indsætte dette DNA-fragment i RMCE-kompatible genomiske locus på grund af dobbeltstrenget samtidig translokation (figur 1). Her, kan kun korrekt RMCE-målrettede kloner gøre lægemiddelresistens takket være en promotor på den indkommende vektor, som genskaber en "fanget," promotor-mindre neomycinresistensgen (NeoR) til stede i R26-genomet af docking-celler (figur 1) 10, 11. Dette resulterer i en meget høj målretning effektivitet, ofte tæt på 100% 11, </ sup> 12. Afslutningsvis RMCE målretning er yderst effektiv og kan anvendes til struktur-funktioner undersøgelser; men det kræver en præfabrikerede genomiske locus.
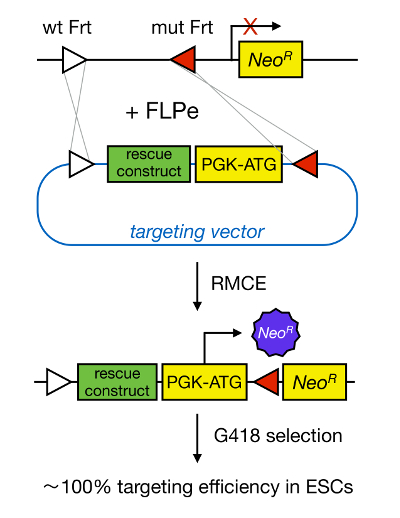
Figur 1. Skematisk repræsentation af RMCE-medieret Målretning. RMCE giver mulighed for udveksling af DNA-segmenter fra et indkommende vektor målretning til en defineret genomisk locus hvis begge harbor to Heterospecifikke FRT-steder (skildret af hvide og røde trekanter). Derudover den konstruerede genomiske locus indeholder et promotorløst og trunkeret neomycin-resistens (NeoR) gen. Ved at tilvejebringe en promotor og startkodon i den indkommende DNA-fragment, kun korrekte rekombinationsbegivenheder genoprette neomycinresistens, hvilket resulterer i effektiviteter høje målretning. Klik her for at se en større version af thans figur.
Genommanipulation i mESCs muliggør frembringelsen af RMCE-kompatibel mus. I 1981, to grupper formået at tage pluripotente celler fra den indre cellemasse (ICM) af blastocyster og opretholdelse dem i kultur 13, 14. mESCs er i stand til selvfornyelse og differentiering i alle typer af embryonale og voksne celler, herunder kim-celle afstamning. Derfor genmålretning i mESCs muliggør omvendt genetiske undersøgelser gennem udvikling af konstitutive eller betingede (ved hjælp af Cre / LoxP-system) KO-mus. Men den klassiske måde at isolere mus ES-celler er meget ineffektiv. Flere store forbedringer har stærkt forøget succesraten for afledt Mesc linjer, herunder anvendelse af et defineret serum-udskiftning (SR) medium 15, vekslende mellem Mesc medium indeholdende SR og føtalt bovint serum (FBS) 16, og anvendelsen af farmakologiske forbindelser, såsom pluripotin eller 2i 17. Pluripotin, en lille syntetisk molekyle, giver mulighed for opformering af mESCs i en udifferentieret tilstand i fravær af leukæmiinhiberende faktor (LIF) og muse embryonale fibroblaster (MEF) 18. Endelig er det blevet vist, at mESCs kan isoleres med en meget høj effektivitet (næsten 100%), når en SR / FBS medium alternerende protokol er kombineret med LIF og pluripotin 19, 20. Disse protokoller muliggør effektiv isolering af RMCE-kompatibel KO mESCs, der efterfølgende kan anvendes til struktur-funktions-undersøgelser.
Dette papir beskriver en fremgangsmåde, der gør det muligt at identificere de vigtigste domæner eller rester i et protein, der er ansvarlige for specifikke cellulære processer. Til dette formål en pipeline af avancerede teknologier, der muliggør effektiv Mesc isolation, målrettede vektor samling, og Mesc målretning var skabed. Som sådan, store paneler med proteinisoformer, domæne mutanter og nedstrømseffektorer kan indføres i KO mESCs og kan evalueres for deres evne til at redde in vitro KO fænotype.
