إنجاز العديد من البروتينات وظيفتها في مجمعات دينامية مع جزيئات أخرى، بوساطة التغيرات كونفورماشونال ورابطات عابرة على مجموعة واسعة من المقاييس الزمنية1،،من23. يقترن بمصدر الطاقة خارجي (مثل ATP) يمكن أن يؤدي إلى اتجاه في دورة وظيفي هذه التفاعلات الدينامية وفي النهاية الحفاظ على ثابت-حالة عدم التوازن في خلية، شرطا أساسيا للحياة.
أجل الفهم الكامل لهذه الآلات الجزيئية، وصف ثابتة يسترشد بدراسات الهيكلية غير كافية. وبالإضافة إلى ذلك، من الضروري الحصول على معرفة طراز الحركية الكامنة وتحديد الثوابت معدل الحركية. العديد من الأساليب القائمة السماح للباحثين بدراسة القوى المحركة للتفاعلات الثنائية بين اثنين من جزيئات الفائدة، مثلاً، السطحية مأكل مثل الطحين الرنين، وأساليب الاسترخاء مع قراءات الطيفية (مثلاً، القفز أو توقف تدفق تقنيات)، والرنين المغناطيسي النووي. ومع ذلك، قابليتها للتطبيق في معظم الحالات يقتصر على أنظمة الدولتين بسيطة (مثلاً، واحد محدد وإحدى الدول غير المنضمة) بسبب متوسط الملازمة للجزء الأكبر من التجارب. في الحالات التي تنطوي على مزيد من الدول أو وسيطة، أنها تعطي فقط خليط معقد من الثوابت معدل. يمكن استرداد طرق جزيء واحد مثل الملقط ضوئية أو مغناطيسية أو اللونين سمفريت و أي جهة مانحة و fluorophore يقبلون واحد، مع عينة المعطل تداولها سطح الثوابت معدل للجميع لاحظ التغييرات كونفورماشونال. ومع ذلك، عندما يتعلق الأمر بالتفاعلات التي تؤثر على موقع ربط واحد أو أكثر، هذه الأساليب لا تزال محدودة والمعلومات المتعلقة بالتفاعلات العلاقة المحتملة للاثنين (أو أكثر) سيكون متاحاً عن طريق الاستنتاجات غير المباشرة من مجموعة من التجارب فقط.
سمفريت متعدد الألوان4،5،،من67،،من89 يتيح الفرصة لدراسة التفاعل بين هذه المكونات مباشرة، في الوقت الحقيقي، وتحت 10من شروط قرب الفسيولوجية. وهذا يسمح أحد للتحقيق على سبيل المثال، ربط تعتمد على تكيف يجند أو البروتين8،،من911. النهج العام للمقدمة هنا لتسمية protein(s) الفائدة في مواقف محددة، تعلق بروتين واحد على سطح الدائرة بالقياس، وتتبع شدة الأسفار على مر الزمن في تيرفم نوع المنشور (للتفاصيل انظر 9 , 12). يمكن تحديده بالقرب المكاني من صبغات مختلفة ثم من نقل الطاقة بين البلدين. تصنيف استراتيجيات تختلف من البروتين للبروتين (استعرض في 13) وتوجد مبادئ توجيهية لتجنب القطع الأثرية في القياسات سمفريت14.
منذ صبغة الجهات مانحة قد نقل الطاقة إلى الأصباغ يقبلون مختلفة في تجربة سمفريت متعدد ألوان، الموضع النسبي لجميع الأصباغ غير قابل للوصول من الإثارة لصبغ واحد وحدها15،16. ولكن في تركيبة مع التناوب الإثارة الليزر (أليكس17، و استعرضت في 18) يوفر هذا الأسلوب جميع المعلومات الزمانية في الثانية الفرعية والقرار الفرعي نانومتر.
في رأس مال، حساب عالية الدقة يمكن أن يتحقق من المعلومات الهيكلية باستخدام المسافات بين صبغ من الجمع بين جميع الأسفار الكثافات في تجربة سمفريت متعدد ألوان مع أليكس. ومع ذلك، نركز هنا على تحديد هوية الدولة والانفصال، فضلا عن استخراج النماذج الحركية، حيث سمفريت متعدد الألوان أمر لا غنى عنه. عندما المطلوب هو “فقط” هيكل التصميم بالتثليث، مجموعة من التجارب سمفريت اللونين أبسط مع نسبة الإشارة إلى الضوضاء عالية يمكن أداء12،19.
ونحن نستخدم fluorescence جزئي (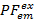 ) كوكيل لنقل الطاقة بين اثنين فلوروفوريس7. PF يحسب من الأسفار كثافة مماثلة لكفاءة الحنق تجربة اللونين:
) كوكيل لنقل الطاقة بين اثنين فلوروفوريس7. PF يحسب من الأسفار كثافة مماثلة لكفاءة الحنق تجربة اللونين:
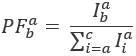
فيها، 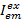 الكثافة في الانبعاثات قناة م بعد الإثارة مع اللون السابق، وهو ج يقبلون مع الطول الموجي الأطول. وتمثل قنوات الكشف عن نفس الموقف في الدائرة عينة ولكن سجل نطاقات طيفية مختلفة من الضوء الفلورية. يتم استخدام نفس المعرف للإثارة والانبعاثات في هذا البروتوكول (أي، “الأزرق”، “الخضراء”، و “أحمر”).
الكثافة في الانبعاثات قناة م بعد الإثارة مع اللون السابق، وهو ج يقبلون مع الطول الموجي الأطول. وتمثل قنوات الكشف عن نفس الموقف في الدائرة عينة ولكن سجل نطاقات طيفية مختلفة من الضوء الفلورية. يتم استخدام نفس المعرف للإثارة والانبعاثات في هذا البروتوكول (أي، “الأزرق”، “الخضراء”، و “أحمر”).
بسبب قصور تجريبية تعتمد كثافة fluorescence يقاس ليس فقط في نقل الطاقة ولكن أيضا على خصائص فلوروفوري والإعداد. بغية الحصول على كفاءة نقل الطاقة الحقيقية بين هذين فلوروفوريس، قد كثافات المقاسة ينبغي تصحيحها. يستند الإجراء التالي على المرجع9. عوامل التصحيح للتسرب الظاهر (lk، أي مكان مخصص للكشف عن الفوتونات من فلوروفوري في قناة لصبغ آخر) وغاما الظاهر (ag، أي العائد الكم الأسفار من الصبغة الكشف عن كفاءة القناة) تم الحصول عليها من آثار جزيء واحد تظهر يقبلون تبيض الحدث.
تسرب الصبغة المانحين في كل قناة يقبلون ممكن يتم حسابها من كافة نقاط البيانات في آثار fluorescence مسجل فيها مقصور صبغ يقبلون لكن الجهة المانحة لا تزال الفلورسنت (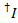 ):
):
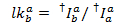
الوسيط للرسم البياني التسرب كعامل التسرب الظاهر. بعد التصحيح للتسرب، يتم تحديد معامل جاما الظاهر من نفس المجموعة من آثار. فإنه يحسب بقسمة تغير الأسفار في قناة يقبلون على التغيير للأسفار في قناة المانحين على تبييض لصبغ يقبلون:
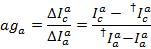
حيث c هو مرة أخرى قناة كشف يقبلون مع الطول الموجي الأطول. الوسيط لتوزيع الناتج كعامل التصحيح الظاهر.
يتم الحصول على كثافة تصحيحها في كل قناة من:
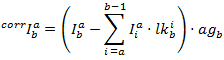
ثم يتم حساب الجبهة الوطنية استناداً إلى:
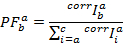
يمكن فصل مختلف قطاعات السكان في الفضاء المتعدد الأبعاد موزعة حسب PFs. الموقف وعرض كل دولة يتحدد باحتواء البيانات مع وظائف الضبابي متعدد الأبعاد. التحسين اللاحقة من هم عالمي واحد استناداً إلى جميع آثار PF يوفر وصفاً كمياً للحركية الملحوظة. حتى التغيرات الصغيرة معدلات قابلة للاكتشاف.
هممس توفر طريقة لاستنتاج نموذج دولة من مجموعة من آثار الوقت صاخبة. هذا النظام يعتبر أن في واحدة من مجموعة منفصلة، الدول المخفية في أي وقت من الأوقات والمراقبة الفعلية (أي الانبعاثات) دالة احتمالية لهذه الدولة الخفية20. في حالة البيانات سمفريت تيرفم، الانبعاثات الاحتمالات بأنا كل دولة وأنا يمكن أن تكون على غرار بوظائف مستمرة الكثافة الاحتمالية الضبابي. في نقاط زمنية منفصلة متباعدة بشكل منتظم، يمكن أن يحدث الانتقال من واحد إلى دولة أخرى وفقا لاحتمال انتقال الثابتة على الوقت ويعتمد فقط على الحالة الراهنة. مصفوفة الانتقال A يحتوي على هذه الاحتمالات الانتقال ij بين جميع الدول المخفية. توزيع الدولة الأولى 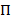 يعطي على الاحتمالات الخاصة بالدولة
يعطي على الاحتمالات الخاصة بالدولة 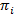 لأول مرة نقطة تتبع الوقت. استخدام نهج الحد الأقصى-احتمال، يمكن تحسين هذه المعلمات لأفضل وصف البيانات مع الأمام-للخلف واوم-ولش خوارزميات20،21. وهذا ينتج المقدرات الحد الأقصى لاحتمال (MLE). وأخيراً، يمكن الاستدلال على تسلسل الدولة التي أنتجت على الأرجح مسار الملاحظات مع خوارزمية Viterbi. خلافا لغيرها من التحليلات هم سمفريت البيانات24،،من25إلى26 نحن لا نستخدم هم كمجرد “تجانس” من البيانات ولكن استخراج نموذج الدولة الحركية من مجموعة البيانات بدون الحاجة لتركيب الوقت يسكن 27من رسوم بيانية. ويتم تحليل هم مع البرامج النصية الداخلية باستخدام برو إيغور. ويستند تنفيذ المدونة مرجع21. نحن توفير مجموعة البرمجيات والبيانات النموذجية على لدينا صفحة ويب كي تتبع القسمين 5 و 6 من هذا البروتوكول (https://www.singlemolecule.uni-freiburg.de/software/3d-fret). البرنامج الكامل متاح عند الطلب.
لأول مرة نقطة تتبع الوقت. استخدام نهج الحد الأقصى-احتمال، يمكن تحسين هذه المعلمات لأفضل وصف البيانات مع الأمام-للخلف واوم-ولش خوارزميات20،21. وهذا ينتج المقدرات الحد الأقصى لاحتمال (MLE). وأخيراً، يمكن الاستدلال على تسلسل الدولة التي أنتجت على الأرجح مسار الملاحظات مع خوارزمية Viterbi. خلافا لغيرها من التحليلات هم سمفريت البيانات24،،من25إلى26 نحن لا نستخدم هم كمجرد “تجانس” من البيانات ولكن استخراج نموذج الدولة الحركية من مجموعة البيانات بدون الحاجة لتركيب الوقت يسكن 27من رسوم بيانية. ويتم تحليل هم مع البرامج النصية الداخلية باستخدام برو إيغور. ويستند تنفيذ المدونة مرجع21. نحن توفير مجموعة البرمجيات والبيانات النموذجية على لدينا صفحة ويب كي تتبع القسمين 5 و 6 من هذا البروتوكول (https://www.singlemolecule.uni-freiburg.de/software/3d-fret). البرنامج الكامل متاح عند الطلب.
الوقت النقاط في البيانات مع الجبهة الوطنية <-1 أو PF > يتم تعيين 2 في أي قناة الكشف عن احتمال انبعاث الحد الأدنى لجميع الدول (10-200). وهذا ما يمنع التحولات المصطنعة في نقاط البيانات هذه.
يتم الحصول على المعلمات لاحتمالات الانبعاثات من تناسب الرسم البياني PF 3D مع وظائف الضبابي كما هو موضح في الخطوة 5، 7. وتحفظ هذه المعلمات ثابتة خلال الاستغلال الأمثل هم.
في النهج الذي قدم، تستخدم مصفوفة الانتقال وناقلات توزيع الدولة الأولى عالمياً لوصف فرقة كاملة من آثار. يتم تحديثها استناداً إلى جميع الجزيئات N من مجموعة البيانات وفقا لمرجعية27.
يتم تحديد معلمات البدء لتوزيع الدولة الأولى من إسقاطات 2D من الرسم البياني PF (الخطوة 5، 3) واحتمالات الانتقال يتم تعيين إلى 0.05 استثناء الاحتمالات البقاء في الدولة نفسها، التي يتم اختيار هذه أن احتمال مغادرة دولة معينة هو تطبيع للوحدة.
يتم استخدام أسلوب تنميط احتمال إعطاء فواصل الثقة (رابطة الدول المستقلة) لكل مرحلة انتقالية معدلات21،22، التي تكون بمثابة تقديرات ذات مغزى لعدم اليقين. لحساب حدود كاريتاس الدولية لمعدل معين، هو ثابت احتمال انتقال الاهتمام إلى قيمة أخرى MLE. وهذا ينتج λ نموذج اختبار ‘. اختبار نسبة (LR) احتمال من احتمالات 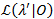 نظراً لمجموعة البيانات تتم 0 استناداً إلى:
نظراً لمجموعة البيانات تتم 0 استناداً إلى:
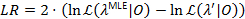
الثقة 95% ملزمة للتوصل إلى المعلمة عندما يتجاوز LR 3.841، كانتيل 95% من س2-التوزيع مع واحد من22،درجة من الحرية23.
هو أظهرت قوة الأسلوب باستخدام في Hsp90. هذا البروتين وفيرة وجدت في البكتريا، وحقيقيات النوى، وهو جزء من استجابة الإجهاد الخلوية28. وهدف المخدرات واعدة في علاج السرطان29. Hsp90 هوموديمير مع جيب ملزم النوكليوتيدات واحدة في المجال الطرفي ن لكل وحدة فرعية30. أنه يمكن أن يخضع للتحولات بين اثنين على الأقل والتشكلات متميزة على الصعيد العالمي، واحدة مغلقة و أحد الطرفي ن تكيف مفتوحة، على شكل V،19،31،32. طبيعة dimeric مباشرة يثير مسألة التفاعل بين هذين الموقعين النوكليوتيدات ملزمة في Hsp90.
في ما يلي، نقدم بروتوكول خطوة بخطوة للحصول على البيانات وتحليلها لتجربة ألوان سمفريت الخميرة Hsp90 والنوكليوتيدات. ربط تعتمد على تكيف PNP أمبير المسمى فلوريسسينتلي (PNP أمبير *، النظير غير هيدروليزابل من ATP) يتم تحليل. تطبيق الإجراء وصف تصاريح دراسة الربط النوكليوتيدات، وفي نفس الوقت التغييرات كونفورماشونال من Hsp90 ومما يكشف عن كوبيراتيفيتي بين اثنين النوكليوتيدات ملزمة جيوب Hsp90.
