Veel eiwitten vervullen hun functie in dynamische complexen met andere moleculen, gemedieerd door conformationele veranderingen en voorbijgaande verenigingen over een brede waaier van tijdschalen1,2,3. Gekoppeld aan een externe energiebron (bijvoorbeeld ATP) deze dynamische interacties kunnen leiden tot directionaliteit in een functionele cyclus en uiteindelijk het onderhouden van de niet-evenwichts steady-state in een cel, een voorwaarde voor het leven.
Om volledig te begrijpen deze moleculaire machines, is een statische beschrijving geleid door structurele studies niet voldoende. Daarnaast is het essentieel te beschikken over kennis van de onderliggende kinetische model en te bepalen van de kinetische snelheidsconstanten. Verschillende bestaande methoden toestaan onderzoekers bestuderen de dynamiek van binaire interacties tussen twee moleculen van belang, bijvoorbeeld oppervlakte plasmon resonantie, ontspanning methoden met een spectroscopische uitlezing (bijvoorbeeld sprong of gestopt-flow technieken), en nucleaire magnetische resonantie. Hun toepasbaarheid is echter in de meeste gevallen beperkt tot eenvoudige tweestaten-systemen (bijvoorbeeld een gebonden en een niet-afhankelijke staat) als gevolg van de gemiddeld die inherent zijn aan bulk experimenten. In gevallen waar meer Staten of tussenproducten zijn betrokken, opleveren zij alleen een complex mengsel van de snelheidsconstanten. Single-molecuul methoden zoals optische of magnetische pincet of twee kleuren smFRET, d.w.z. een donor en één acceptor fluorophore, met een oppervlak-geïmmobiliseerd steekproef kunnen de snelheidsconstanten herstellen voor alle conformationele wijzigingen waargenomen. Echter wanneer het gaat om interacties op het gebied van meer dan één binding site, deze methoden beperkt blijven en de informatie over de wisselwerkingen mogelijk correlatie van de twee (of meer) zal alleen bereikbaar zijn via indirecte conclusies uit een aantal experimenten.
Multi kleur smFRET4,5,6,7,8,9 biedt de mogelijkheid te bestuderen van de interactie tussen deze componenten direct, in real-time en onder in de buurt van de fysiologische omstandigheden10. Hierdoor kan een te onderzoeken bijvoorbeeld de conformatieafhankelijke binding van een ligand of een ander eiwit8,–9,11. De hier gepresenteerde algemene benadering is om de expressie gebrachte eiwitten van belang op specifieke posities, één eiwit hechten aan het oppervlak van de meting-kamer, en het bijhouden van de intensiteit van de fluorescentie na verloop van tijd op een prisma-type TIRFM (voor details zie 9 label , 12). de ruimtelijke nabijheid van de verschillende kleurstoffen kan vervolgens worden bepaald uit de energie-overdracht tussen hen. Labelen van strategieën kan variëren van eiwit tot proteïne (herzien in 13) en richtsnoeren om te voorkomen dat artefacten in smFRET metingen bestaan14.
Aangezien de kleurstof van een donor energie in verschillende acceptor kleurstoffen in een multi kleur smFRET experiment overbrengen kan, is de relatieve positie van alle kleurstoffen niet toegankelijk vanaf excitatie van een kleurstof alleen15,16. Maar deze methode biedt in combinatie met afwisselende laser excitatie (ALEX17en gerecenseerd in 18) alle spatio-temporele informatie bij sub tweede en sub nanometer resolutie.
In principe, hoge resolutie structurele informatie kan worden bereikt door gebruik te maken van de afstanden tussen kleurstof berekend op basis van de combinatie van alle fluorescentie intensiteiten in een multi kleur smFRET experiment met ALEX. Echter richten hier we ons op staat identificatie en scheiding, evenals de winning van kinetische modellen, waar meerdere kleuren smFRET onontbeerlijk. Wanneer “alleen” structuurbepaling door triangulatie is gewenst, kunnen een aantal eenvoudiger twee kleuren smFRET experimenten met hoge signaal-/ ruisverhouding uitgevoerde12,19.
We gebruiken de gedeeltelijke fluorescentie (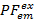 ) als een proxy voor de energie-overdracht tussen twee fluorophores7. De PF is berekend op basis van de fluorescentie intensiteit analoog aan de efficiëntie van de FRET van een experiment van twee kleuren:
) als een proxy voor de energie-overdracht tussen twee fluorophores7. De PF is berekend op basis van de fluorescentie intensiteit analoog aan de efficiëntie van de FRET van een experiment van twee kleuren:
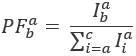
Waar, 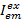 is de intensiteit in emissie kanaal em na excitatie met kleur ex, en c is de acceptor met de langste golflengte. Detectie kanalen vertegenwoordigen dezelfde positie in de monsterkamer maar record verschillende spectrale reeksen van de fluorescentie-licht. Dezelfde id voor excitatie en emissie in dit protocol worden gebruikt (dat wil zeggen, “blauw”, “groene” en “rode”).
is de intensiteit in emissie kanaal em na excitatie met kleur ex, en c is de acceptor met de langste golflengte. Detectie kanalen vertegenwoordigen dezelfde positie in de monsterkamer maar record verschillende spectrale reeksen van de fluorescentie-licht. Dezelfde id voor excitatie en emissie in dit protocol worden gebruikt (dat wil zeggen, “blauw”, “groene” en “rode”).
De gemeten fluorescentie-intensiteiten hangen vanwege experimentele tekortkomingen van niet alleen op de energie-overdracht, maar ook op fluorophore en setup-eigenschappen. Met het oog op het spuitrendement waar energie tussen twee fluorophores, de gemeten intensiteiten moeten worden gecorrigeerd. De volgende procedure is gebaseerd op referentie9. Correctiefactoren voor zichtbare lekkage (lk, dat wil zeggen, de detectie van fotonen van een fluorophore in een kanaal voor een andere kleurstof aangewezen) en schijnbare gamma (ag, dat wil zeggen, de fluorescentie quantumrendement van de kleurstof en de de efficiëntie van de detectie van het kanaal) single-molecuul sporen die aantonen dat een accepteerder bleken gebeurtenis worden verkregen.
De lekkage van de kleurstof van de donor in elk mogelijk acceptor kanaal is berekend op basis van alle gegevenspunten in de sporen van de opgenomen fluorescentie waar de acceptor kleurstof gebleekt maar de donor is nog steeds TL (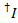 ):
):
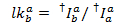
De mediaan van de lekkage histogram wordt gebruikt als de schijnbare lekkage factor. Na correctie voor lekkage, wordt de schijnbare gamma factor bepaald door de dezelfde set van sporen. Het wordt berekend door de verandering van fluorescentie in de acceptor kanaal door de verandering van fluorescentie in het kanaal van de donor bij het bleken van de acceptor kleurstof:
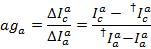
Waarbij c staat weer voor de detectie-kanaal voor de acceptor met de langste golflengte. De mediaan van de resulterende verdeling wordt gebruikt als de schijnbare correctiefactor.
De gecorrigeerde intensiteiten in elk kanaal worden verkregen door:
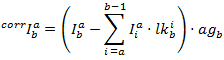
De PF wordt dan berekend volgens:
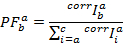
Verschillende bevolkingsgroepen kunnen worden gescheiden in de multi-dimensionale ruimte overspannen door de PF-s. De positie en de breedte van elke staat wordt bepaald door het aanbrengen van de gegevens met multi-dimensionale Gaussian functies. Latere optimalisatie van één wereldwijde HMM gebaseerd op alle sporen van de PF biedt een kwantitatieve beschrijving van de waargenomen kinetiek. Zelfs kleine veranderingen van de tarieven zijn waarneembaar.
HMMs bieden een manier van afgeleid van een staatsmodel uit een collectie van luidruchtige tijd sporen. Het systeem wordt beschouwd als in één van een reeks van discrete, verborgen frames op een bepaald moment en de eigenlijke observatie (dat wil zeggen, de emissie) is een probabilistische functie van deze verborgen staat20. In het geval van TIRFM smFRET gegevens, kan de emissie waarschijnlijkheden bik per staat ik worden gemodelleerd door continue Gaussian waarschijnlijkheid dichtheid functies. Bij regelmatig verdeelde discrete tijd punten, kunnen overgangen van de ene naar een andere staat optreden volgens de overgang waarschijnlijkheid dat tijd-invariant is en hangt alleen af van de huidige status. De overgang matrix A bevat deze overgang waarschijnlijkheden eenij tussen alle verborgen frames. De verdeling van de begintoestand 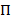 geeft de staatspecifieke waarschijnlijkheid
geeft de staatspecifieke waarschijnlijkheid 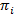 voor het eerste punt van de tijd van een spoor van de tijd. Met behulp van een maximaal-kans aanpak, kunnen deze parameters worden geoptimaliseerd om het best beschrijven de gegevens met de vooruit-achteruit en Baum-Welch algoritmen20,21. Dit levert de maximale waarschijnlijkheid schatters (MLE). Ten slotte kan de volgorde van de staat die waarschijnlijk geproduceerd het traject van opmerkingen met de Viterbi algoritme worden afgeleid. In tegenstelling tot andere HMM analyses van smFRET gegevens24,25,26 wij gebruiken niet de HMM als een louter “glad” van de gegevens, maar de kinetische staatsmodel uittreksel uit de gegevensset zonder de noodzaak voor het aanbrengen van de nadruktijd histogrammen27. HMM analyse wordt gedaan met in-house scripts met Igor Pro. Tenuitvoerlegging van de gedragscode is gebaseerd op referentie21. Wij bieden een software kit en voorbeeldige gegevens op onze website om te volgen van de punten 5 en 6 van dit protocol (https://www.singlemolecule.uni-freiburg.de/software/3d-fret). Volledige software is beschikbaar op aanvraag.
voor het eerste punt van de tijd van een spoor van de tijd. Met behulp van een maximaal-kans aanpak, kunnen deze parameters worden geoptimaliseerd om het best beschrijven de gegevens met de vooruit-achteruit en Baum-Welch algoritmen20,21. Dit levert de maximale waarschijnlijkheid schatters (MLE). Ten slotte kan de volgorde van de staat die waarschijnlijk geproduceerd het traject van opmerkingen met de Viterbi algoritme worden afgeleid. In tegenstelling tot andere HMM analyses van smFRET gegevens24,25,26 wij gebruiken niet de HMM als een louter “glad” van de gegevens, maar de kinetische staatsmodel uittreksel uit de gegevensset zonder de noodzaak voor het aanbrengen van de nadruktijd histogrammen27. HMM analyse wordt gedaan met in-house scripts met Igor Pro. Tenuitvoerlegging van de gedragscode is gebaseerd op referentie21. Wij bieden een software kit en voorbeeldige gegevens op onze website om te volgen van de punten 5 en 6 van dit protocol (https://www.singlemolecule.uni-freiburg.de/software/3d-fret). Volledige software is beschikbaar op aanvraag.
Tijd van punten in het gegevens met PF < -1 of PF > 2 in elk kanaal detectie de minimale uitstoot kans voor alle Staten (10-200) zijn toegewezen. Hiermee voorkomt u dat kunstmatige overgangen op deze gegevenspunten.
De parameters voor de emissie waarschijnlijkheid worden verkregen van de pasvorm van de 3D histogram van de PF met Gaussiaans functies zoals beschreven in stap 5.7. Deze parameters worden vast gehouden tijdens de optimalisatie van de HMM.
In de gepresenteerde benadering, zijn de begintoestand distributie vector en de transitie matrix wereldwijd gebruikt om te beschrijven het hele ensemble van sporen. Zij zijn bijgewerkt op basis van alle N -moleculen van de gegevensset volgens referentie27.
Startparameters voor de distributie van de oorspronkelijke toestand worden bepaald uit 2D projecties van de PF -histogram (stap 5.3) en de waarschijnlijkheid van de overgang zijn ingesteld op 0.05 met uitzondering van de waarschijnlijkheid om te verblijven in dezelfde staat, die zodanig zijn gekozen dat is de kans op een bepaalde toestand laten genormaliseerd naar eenheid.
Een waarschijnlijkheid profiling methode wordt gebruikt om betrouwbaarheidsintervallen (CIs) voor alle overgang tarieven21,22, die dienen als zinvolle schattingen voor hun onzekerheid. Voor de berekening van de grenzen van de CI voor een bepaald tempo, is de kans op overgang van belang op een andere waarde dan de MLE bevestigd. Dit levert de test model λ’. Een waarschijnlijkheid verhouding (LR) test van de kans op 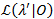 gezien de gegevensset 0 wordt uitgevoerd volgens:
gezien de gegevensset 0 wordt uitgevoerd volgens:
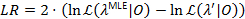
De betrouwbaarheid van 95% gebonden voor de parameter wordt bereikt wanneer LR 3.841, de 95% kwantiel van een x2 overschrijdt-verdeling met een mate van vrijheid22,23.
De kracht van de methode is aangetoond door middel van de Hsp90. Deze overvloedige eiwit is gevonden in bacteriën en eukaryoten, en maakt deel uit van de cellulaire stress reactie28. Het is een veelbelovende drug doelwit van kanker behandeling29. Hsp90 is een homodimer met één nucleotide-bindende zak op het N-terminaal gebied van elke subeenheid30. Het kunt overgangen tussen ten minste twee wereldwijd verschillende conformaties, één gesloten en één N-terminal open, V-vormige conformatie19,31,32ondergaan. De dimeric aard vraag direct de van de wisselwerking tussen de twee nucleotide bandplaatsen in Hsp90.
In het volgende, bieden wij een stapsgewijze protocol voor de data-acquisitie en de analyse van een experiment van de drie kleuren smFRET op gist Hsp90 en nucleotide. De conformatieafhankelijke binding van fluorescently geëtiketteerde AMP-PNP (AMP-PNP *, een niet-hydrolyseerbare analoog van ATP) wordt geanalyseerd. De toepassing van de beschreven procedure toelaat de studie van de nucleotide-bindende en tegelijkertijd de conformationele veranderingen van Hsp90 en daardoor onthult de Allosterie tussen de twee nucleotide-bindende zakken van Hsp90.
