Многие белки выполняют свои функции в динамической комплексы с другими молекулами, посредничестве конформационные изменения и переходных ассоциаций по широкому спектру сроков1,2,3. В сочетании с внешнего источника энергии (например, АТФ) эти динамические взаимодействия может привести к получателю в цикле функциональных и в конечном счете сохранить неравновесного установившемся в клетке, необходимым условием для жизни.
Для того чтобы полностью понять эти молекулярные машины, статическое описание, руководствуясь структурных исследований не является достаточным. Кроме того важно иметь знания о базовой кинетической модели и определить константы кинетической скорости. Несколько существующих методов позволяют исследователям изучить динамику бинарных взаимодействий между двумя молекулами интерес, например, поверхностного плазмон резонанс, методы релаксации с спектроскопических индикация (например, прыгать или остановить поток методы) и ядерного магнитного резонанса. Однако их применимости, в большинстве случаев, ограничивается простых систем двух государств (например, одна граница и одно государство, несвязанных) благодаря усреднение присущие массовых экспериментов. В тех случаях, когда речь идет о более государств или промежуточных продуктов они дают только сложная смесь константы скорости. Одноместный молекула методы, такие как магнитные или оптические пинцеты или два цвета smFRET, то есть, один из доноров и один акцепторной Флюорофор, с образец поверхности прикол может восстановить константы скорости для всех наблюдаемых конформационных изменений. Однако когда дело доходит до взаимодействия, затрагивающих более одной привязки сайта, эти методы по-прежнему ограничены, и информация о возможной корреляции двух (или более) взаимодействия только будут доступны через косвенные выводы из набора экспериментов.
Многоцветные smFRET4,5,6,,78,9 предлагает возможность изучить взаимодействие между этими компонентами непосредственно в режиме реального времени и под рядом физиологические условия10. Это позволяет исследовать например, конформация зависимых Связывание лиганда или другой белок8,9,11. Общий подход, представленные здесь является маркировать белки интерес в определенных позициях, прикрепить один белка на поверхности измерительной камеры, а также отслеживать интенсивности флуоресценции со временем на призму типа TIRFM (для подробности см 9 , 12). пространственная близость различных красителей, затем может быть определено из передачи энергии между ними. Маркировка стратегии может варьироваться от белка белка (обзор в 13) и руководящие принципы, чтобы избежать артефактов в smFRET измерения существуют14.
Поскольку краска доноров может передавать энергию различных акцептора красителей в эксперименте многоцветные smFRET, относительное положение всех красителей не доступен от возбуждения одного красителя только15,16. Но в сочетании с чередующимися лазерного возбуждения (Алекс17и рассмотренных в 18) Этот метод обеспечивает все пространственно временной информации на секунды и резолюции Подкомиссии нанометров.
В принципе высоким разрешением, которую структурная информация может быть достигнуто с помощью расстояния между краситель рассчитывается из комбинации всех интенсивностью флюоресценции в многоцветные smFRET эксперимент с АЛЕКСОМ. Однако здесь мы сосредоточиться на идентификации состояний и разделения, а также добыча кинетической модели, где smFRET многоцветные незаменим. Когда «только» для определения структуры путем триангуляции, набор проще-двухцветный smFRET экспериментов с высоким соотношением сигнал шум может быть осуществляется12,19.
Мы используем частичное флюоресценция (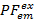 ) как прокси для передачи энергии между двумя флуорофоров7. PF рассчитывается от интенсивности флуоресценции аналогична эффективности ладу-двухцветный эксперимента:
) как прокси для передачи энергии между двумя флуорофоров7. PF рассчитывается от интенсивности флуоресценции аналогична эффективности ладу-двухцветный эксперимента:
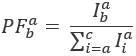
Где 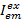 интенсивность выбросов канал ет после возбуждения с цветом ex, и c акцептор с длинной волны. Обнаружение каналов представляют собой ту же позицию в пример камеры, но записи различных спектральных диапазонах света флуоресцирования. Тот же идентификатор для возбуждения и выбросов используются в настоящем Протоколе (т.е., «синий», «зеленых» и «красных»).
интенсивность выбросов канал ет после возбуждения с цветом ex, и c акцептор с длинной волны. Обнаружение каналов представляют собой ту же позицию в пример камеры, но записи различных спектральных диапазонах света флуоресцирования. Тот же идентификатор для возбуждения и выбросов используются в настоящем Протоколе (т.е., «синий», «зеленых» и «красных»).
Из-за экспериментальной недостатки интенсивностью флюоресценции измеренных зависит не только на передачу энергии, но и на Флюорофор и установки свойств. Чтобы получить истинное энергетической эффективности передачи между двумя флуорофоров, измеряемых интенсивностей должны быть исправлены. Следующая процедура основана на ссылку9. Поправочные коэффициенты для явного утечки (lk, т.е. обнаружение фотонов от Флюорофор в канале, предназначенные для другого красителя) и очевидной гамма (ag, т.е. квантовый выход флуоресценции красителя и эффективность обнаружения канала) получаются из одной молекулы следы, которые показывают акцептора отбеливания событие.
Утечки доноров красителя в каждый канал возможно акцептора рассчитывается от всех точек данных в записанных флуоресценции следы где краска акцептора отбеленная, но доноров до сих пор флуоресцентные (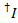 ):
):
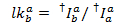
Медиана утечки гистограммы используется как фактор явно утечки. После коррекции для утечки очевидной гамма фактор определяется из того же набора трассировок. Она рассчитывается путем деления изменения флюоресценции в акцепторной канал изменения флюоресценции в канале доноров после отбеливания акцептора красителя:
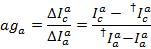
Где c снова является каналом обнаружения акцептор с длинной волны. Медиана распределения результирующей используется как очевидной поправочный коэффициент.
Исправленные интенсивностей в каждом канале получаются путем:
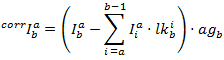
PF рассчитывается согласно:
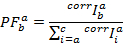
Различные группы населения могут быть разделены в многомерном пространстве, занимаемых PFs. Положение и ширину каждого государства определяется путем установки данных с многомерной функции Гаусса. Последующей оптимизации одной глобальной Хм, основанные на все следы PF обеспечивает количественное описание наблюдаемых кинетики. Даже небольшие изменения ставок обнаруживаются.
HMMs обеспечивают способ выведение модель государства из коллекции шумных следы. Система считается одним из набора дискретных, скрытые состояния в любой момент времени и фактические наблюдения (т.е. выбросов) является вероятностное функцией этой скрытой государственной20. В случае TIRFM smFRET данных выбросов вероятностей b,я за государство, я могут быть смоделированы путем непрерывного Гауссова плотность вероятности функций. В точках через равные интервалы времени дискретных переходы от одного до другого государства может произойти согласно переход вероятность того, что время инвариантная и зависит только от текущего состояния. Матрицы перехода A содержит эти вероятности перехода ij между всеми скрытые государствами. Начальное состояние распространения 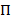 дает вероятностей положени специфически
дает вероятностей положени специфически 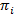 для первой точки время время трассировки. Методом максимального правдоподобия, эти параметры могут быть оптимизированы, чтобы лучше всего описать данные с вперед-назад и Баум-Уэлч алгоритмов20,21. Это дает оценок максимального правдоподобия (MLE). Наконец государство последовательность, которая скорее производится траектории наблюдений может быть выведено с Витерби алгоритм. В отличие от других анализов Хм smFRET данных24,,2526 мы не используем HMM как всего лишь «разглаживание» данных но экстракт кинетическая общегосударственной модели из набора данных без необходимости для установки времени задержки гистограммы27. HMM анализ делается с собственных скриптов с использованием Игорь Pro. Реализация кода основана на ссылку21. Мы предлагаем комплект программного обеспечения и образцовые данные на нашем сайте для того, чтобы следовать разделы 5 и 6 настоящего Протокола (https://www.singlemolecule.uni-freiburg.de/software/3d-fret). Полное программное обеспечение предоставляется по запросу.
для первой точки время время трассировки. Методом максимального правдоподобия, эти параметры могут быть оптимизированы, чтобы лучше всего описать данные с вперед-назад и Баум-Уэлч алгоритмов20,21. Это дает оценок максимального правдоподобия (MLE). Наконец государство последовательность, которая скорее производится траектории наблюдений может быть выведено с Витерби алгоритм. В отличие от других анализов Хм smFRET данных24,,2526 мы не используем HMM как всего лишь «разглаживание» данных но экстракт кинетическая общегосударственной модели из набора данных без необходимости для установки времени задержки гистограммы27. HMM анализ делается с собственных скриптов с использованием Игорь Pro. Реализация кода основана на ссылку21. Мы предлагаем комплект программного обеспечения и образцовые данные на нашем сайте для того, чтобы следовать разделы 5 и 6 настоящего Протокола (https://www.singlemolecule.uni-freiburg.de/software/3d-fret). Полное программное обеспечение предоставляется по запросу.
Времени точек данных с PF < -1 или PF > 2 в любой канал обнаружения назначаются минимальные выбросы вероятность для всех государств (10-200). Это предотвращает искусственных переходы в этих точках данных.
Параметры для вероятностей выбросов получаются из fit 3D PF гистограммы с гауссова функций, как описано в шаге 5.7. Эти параметры хранятся фиксированной во время оптимизации Хм.
В представленный подход начальное состояние распределение вектора и матрицы перехода используются глобально описать весь ансамбль следов. Они обновляются на основе всех N молекул из набора данных по ссылке27.
Параметры запуска для распространения начального состояния определяются из 2D прогнозы PF гистограммы (шаг 5.3) и вероятности перехода устанавливаются 0,05 за исключением вероятность остаться в том же состоянии, которые выбираются такие что вероятность оставить определенного состояния нормализуется к единству.
Метод профилирования вероятность используется дать доверительные интервалы (ди) для всех перехода ставки21,22, которые служат в качестве значимых оценок их неопределенности. Для вычисления границы CI для конкретной скорости, вероятность перехода интерес фиксируется значение, отличное от ОМП. Это дает тест модель λ’. Соотношение (LR) тест вероятности вероятности 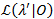 набора данных 0 выполняется согласно:
набора данных 0 выполняется согласно:
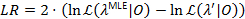
95% доверия, предназначенной для параметра достигается, когда LR превышает 3.841, 95% квантиль о x2-распределение с одной степенью свободы22,23.
Сила метода подтверждается с помощью Hsp90. Этот белок в изобилии встречается в бактерий и эукариот и является частью клеточного стресса ответ28. Это перспективные цели наркотиков в рака лечение29. Hsp90 является Антуану с один карман привязки нуклеотидов в N-концевой домен каждая субъединица30. Он может подвергнуться переходы между по крайней мере двух глобально собственный конформации, закрывается и один N-терминала открытым, V-образный конформации19,31,32. Димерной природы непосредственно поднимает вопрос о взаимосвязи между двумя сайтами привязки нуклеотидов в Hsp90.
В следующем, мы предоставляем пошаговые протокол для сбора и анализа данных эксперимента трехцветной smFRET дрожжи Hsp90 и нуклеотидов. Конформации зависимые привязки дневно обозначенные AMP-PNP (AMP-PNP *, не гидролизуется аналоговый АТФ) анализируется. Приложение описанная процедура позволяет исследование нуклеотида привязки и в то же время конформационные изменения Hsp90 и таким образом раскрывает кооперативность между двумя нуклеотидов привязки очагов Hsp90.
