许多蛋白质完成他们的作用在动态复合体与其他分子, 由构象变化和瞬态协会介导在宽广的范围的时间表1,2,3。耦合到外部能量源 (例如, ATP) 这些动态交互作用可以在功能循环中导致方向性, 并最终维持单元格中的非平衡稳定状态, 这是生命的先决条件。
为了充分理解这些分子机器, 以结构研究为指导的静态描述是不够的。此外, 重要的是要了解基本的动力学模型和确定的动力学速率常数。一些现有的方法让研究人员研究两个感兴趣的分子之间的二元相互作用的动力学,例如,表面等离子体共振, 具有光谱读数的松弛方法 (例如,跳转或停止流技术), 和核磁共振。但是, 它们的适用性在大多数情况下仅限于简单的双状态系统 (例如,一个绑定和一个未绑定状态), 这是由于批量实验所固有的平均值。在涉及更多的状态或中间体的情况下, 它们只产生速率常数的复杂混合。单分子方法, 如光学或磁性镊子或双色 smFRET,即,一个施主和一个受体荧光, 与表面固定的样品可以恢复所有观察到的构象变化的速率常数。但是, 当涉及到影响多个绑定站点的交互时, 这些方法仍然是有限的, 而关于两个 (或多个) 交互的可能相关性的信息只能通过一组实验的间接结论访问。
多色 smFRET4,5,6,7,8,9提供了一个机会, 直接研究这些组件之间的交互, 在实时和下接近生理情况10。这允许一个调查例如, 一个配体或另一个蛋白质的构象依赖性的捆绑8,9,11。这里介绍的总体方法是在特定位置标记感兴趣的蛋白质, 将一个蛋白质附加到测量室的表面, 并在棱镜型显微镜上跟踪时间的荧光强度 (详见9,12). 不同染料的空间接近度可以通过它们之间的能量转移来确定。标记策略可能因蛋白质而异 (在13中进行了审查), 并且避免了 smFRET 测量中的工件存在14的准则。
由于施主染料可能转移能量对不同的接受者染料在多颜色 smFRET 实验, 所有染料的相对位置不是可接近的从励磁一个染料单独15,16。但与交替激光励磁 (亚历克斯17, 并在18中进行了审查) 结合使用, 该方法提供了所有的时空信息, 分二次和亚纳米分辨率。
在主, 高分辨率结构信息可以通过使用的染料之间的距离计算, 从所有荧光强度的组合在一个多色 smFRET 实验与亚历克斯。然而, 在这里, 我们的重点是状态识别和分离以及动力学模型的提取, 其中多色 smFRET 是必不可少的。当 “只” 的结构确定由三角化是需要的, 一组简单的双色 smFRET 实验的高信噪比可以执行12,19。
我们使用部分荧光 () 作为两个荧光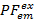 7之间能量传输的代理。PF是从与双色实验的烦恼效率类似的荧光强度计算得出的:
7之间能量传输的代理。PF是从与双色实验的烦恼效率类似的荧光强度计算得出的:
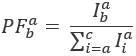
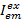 其中, 是在发射通道上的强度与颜色ex之后的em , 而c是波长最长的受体。检测通道在样品室中代表同一位置, 但记录荧光光的不同光谱范围。此协议中使用了相同的激发和发射标识符 (即“蓝色”、”绿色” 和 “红色”)。
其中, 是在发射通道上的强度与颜色ex之后的em , 而c是波长最长的受体。检测通道在样品室中代表同一位置, 但记录荧光光的不同光谱范围。此协议中使用了相同的激发和发射标识符 (即“蓝色”、”绿色” 和 “红色”)。
由于实验的缺陷, 所测得的荧光强度不仅取决于能量的转移, 而且还依赖于荧光和设置的性质。为了获得两个荧光之间的真实能量传递效率, 必须对测量的强度进行修正。以下过程基于引用9。明显渗漏的校正因子 (lk,即从指定给另一染料的通道中的荧光中检测光子) 和表观伽玛 (ag,即,染料的荧光量子屈服和该通道的检测效率是从单分子的痕迹, 显示一个受体漂白事件。
在所有可能的受体通道上的施主染料的泄漏是从记录的荧光痕迹的所有数据点计算的, 那里的受体染料漂白, 但施主仍然荧光 ():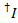
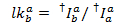
将泄漏直方图的中值作为表观泄漏系数。在对泄漏进行校正后, 视伽玛系数是从同一组跟踪中确定的。它是通过将受体通道中的荧光变化除以施主通道中的发光变化来计算受体染料的漂白:
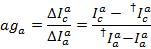
其中, c再次是最长波长的接受方的检测通道。所产生的分布的中位数被用作表观校正因子。
每个通道的校正强度通过以下方法获得:
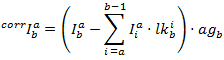
然后根据以下内容计算PF :
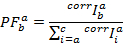
在PF所跨越的多维空间中, 可以分隔不同的填充。用多维高斯函数拟合数据, 确定了各状态的位置和宽度。基于所有PF跟踪的一个全局 HMM 的后续优化提供了观察到的动力学的定量描述。即使是微小的变化率是可察觉的。
模型提供一种从嘈杂的时间痕迹集合中推断状态模型的方法。系统被认为是在任意给定时间的一组离散的隐藏状态中的一个, 而实际观测 (即发射) 是这个隐藏状态的概率函数20。在显微镜 smFRET 数据的情况下, 可通过连续高斯概率密度函数对每个状态i的发射概率bi 进行建模。在通常间隔的离散时间点, 从一个到另一个状态的转换可以根据时间不变的过渡概率发生, 并且只取决于当前状态。转换矩阵a包含这些转换概率aij 在所有隐藏状态之间。初始状态分布为时间跟踪的第一个时间点提供特定于状态的概率. 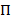
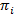 使用最大似然方法, 这些参数可以优化, 以最有效地描述数据的前向向后和鲍姆-韦尔奇算法20,21。这就产生了最大似然估计量。最后, 最有可能产生观测轨迹的状态序列可以用维特比算法来推断。与 smFRET 数据的其他 hmm 分析相反,24、2526我们不使用 hmm 作为数据的 “平滑”, 而是从数据集中提取动态状态模型, 而不需要拟合驻留时间直方图27。HMM 分析是做内部脚本使用伊戈尔 Pro。代码的实现基于引用21。我们在我们的网页上提供一个软件套件和模范数据, 以遵循本协议 (https://www.singlemolecule.uni-freiburg.de/software/3d-fret) 的5和6节。可根据要求提供完整的软件。
使用最大似然方法, 这些参数可以优化, 以最有效地描述数据的前向向后和鲍姆-韦尔奇算法20,21。这就产生了最大似然估计量。最后, 最有可能产生观测轨迹的状态序列可以用维特比算法来推断。与 smFRET 数据的其他 hmm 分析相反,24、2526我们不使用 hmm 作为数据的 “平滑”, 而是从数据集中提取动态状态模型, 而不需要拟合驻留时间直方图27。HMM 分析是做内部脚本使用伊戈尔 Pro。代码的实现基于引用21。我们在我们的网页上提供一个软件套件和模范数据, 以遵循本协议 (https://www.singlemolecule.uni-freiburg.de/software/3d-fret) 的5和6节。可根据要求提供完整的软件。
在任何检测通道中, pf <-1 或pf > 2 的数据中的时间点被分配给所有状态 (10-200) 的最小发射概率。这可以防止在这些数据点进行人工转换。
根据5.7 步中描述的高斯函数的 3D PF直方图的拟合, 得到了发射概率的参数。这些参数在优化 HMM 的过程中保持固定。
在所提出的方法中, 初始状态分布向量和转换矩阵在全局上用于描述整个跟踪的集合。它们根据引用27的数据集中的所有N分子进行更新。
初始状态分布的起始参数由PF直方图的2D 投影确定 (步骤 5.3), 而转换概率设置为 0.05, 但概率保持在同一状态, 这是选择离开某一状态的概率被归为统一。
一种似然分析方法用于给所有过渡速率的置信区间 (CIs)21,22, 它作为对其不确定性的有意义的估计。为了计算某一特定速率下的 CI 的边界, 将利率的转移概率定为非极大似然的值。这将生成测试模型λ “。给定数据集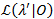 0的可能性的可能性比 (LR) 测试是根据以下内容执行的:
0的可能性的可能性比 (LR) 测试是根据以下内容执行的:
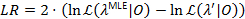
当LR超过 3.841, 95% 位的x2-分布与一个自由度22,23时, 参数的置信度达到了95%。
使用 Hsp90 演示了该方法的威力。这种丰富的蛋白质是在细菌和真核生物中发现的, 是细胞应激反应的一部分28。它是一个有前途的药物靶在癌症治疗29。Hsp90 是在每个子单元30的 N 终端域中具有一个核苷酸绑定口袋的 homodimer。它可以在至少两个全局不同的构象之间进行转换, 一个闭合和一个 N 终端开放, V 形构象19,31,32。二性质直接引起了 Hsp90 两个核苷酸结合位点之间相互作用的问题。
在下面的过程中, 我们提供了一个分步协议的数据获取和分析三色 smFRET 实验酵母 Hsp90 和核苷酸。分析了荧光标记的安培-pnp (amp)、非水解模拟 ATP 的构象依赖性结合。被描述的做法的应用允许研究核苷酸结合和同时 Hsp90 的构象变动和从而显露协同在二个核苷酸结合的口袋 Hsp90 之间。
