تفي الميتوكوندريا بتوفير الطاقة الرئيسي وهي عضية مجزأة تساهم في عمليات الطاقة الحيوية والتمثيل الغذائي الخلوية الأساسية مثل ابتنائية النيوكليوتيدات والدهون والأحماض الأمينية والتكوين الحيوي لعنقود الحديد والكبريت وتتورط في إشارات مثل موت الخلايا المتحكم فيه1،2،3 . تساهم الطاقة الحيوية للميتوكوندريا من خلال الفسفرة التأكسدية في جميع العمليات الخلوية داخل الخلية تقريبا ، وبالتالي ، ترتبط اختلالات الميتوكوندريا ذات الأصل الأولي أو الثانوي بمجموعة واسعة من الحالات المرضية 4,5. خلل الميتوكوندريا لا ينطوي فقط على تغييرات في البنية أو كثافة الميتوكوندريا ولكن أيضا في جودة وتنظيم الجهاز التنفسي6. يشمل هذا العنصر النوعي التحكم في الركيزة ، وخصائص الاقتران ، والتعديلات اللاحقة للترجمة ، وديناميكيات cristae ، والمجمعات الفائقة التنفسية 7,8. لذلك ، فإن التحليل الدقيق للطاقة الحيوية الميتوكوندريا للنهج التجريبية والتشخيصية لتقييم استقلاب الطاقة للخلية مهم في الصحة والمرض.
الفسفرة التأكسدية للميتوكوندريا (OXPHOS) هي سلسلة من التفاعلات داخل الجهاز التنفسي أو نظام نقل الإلكترون (ETS) لتوليد الطاقة الخلوية من خلال الأدينوسين ثلاثي الفوسفات (ATP)9. تولد الخطوة متعددة الأنزيمات لتسخير الطاقة من تدفق الإلكترون عبر المجمعين الأول والثاني إلى المركب الرابع تدرج بروتون كهروكيميائي عبر غشاء الميتوكوندريا الداخلي ، يستخدم لاحقا لفسفرة ثنائي فوسفات الأدينوسين (ADP) إلى ATP عبر المركب V (F1FO ATP SYNTHASE) (الشكل 1A).
أولا ، يتم توليد حاملات ثنائية الإلكترونات خلال دورة ثلاثي الكربوكسيل (TCA) ، تحلل السكر ، وأكسدة البيروفات: نيكوتيناميد الأدينين ثنائي النوكليوتيد (NADH) وثنائي هيدروفلافين الأدينين ثنائي النوكليوتيد (FADH2). يتأكسد NADH في المركب I (نازعة هيدروجيناز NADH) ، حيث يتم نقل إلكترونين إلى الإنزيم المساعد Q (يتم تقليل الكينون إلى الكينول) ، بينما يتم ضخ البروتونات في الفضاء بين الأغشية (IMS). ثانيا ، المركب II (نازعة هيدروجيناز سكسينات) يؤكسد FADH2 ويغذي الإلكترونات إلى الإنزيم المساعد Q دون ضخ البروتونات. ثالثا ، في المركب الثالث (السيتوكروم ج أوكسيدوريدوكتاز) ، يتم نقل الإلكترونات من الإنزيم المساعد Q إلى السيتوكروم c بينما يتم ضخ البروتونات في IMS. رابعا ، ينقل السيتوكروم c الإلكترونات إلى المركب IV (Cytochrome c oxidase) ، وهو المركب الأخير لضخ البروتونات ، وحيث يعمل الأكسجين كمستقبل للإلكترون لاستيعاب البروتونات ، مما يشكل الماء في النهاية. هذا هو الأكسجين الذي تستهلكه الميتوكوندريا والذي يمكن قياسه بواسطة oxygraph. وأخيرا ، يتم استخدام البروتونات المتولدة من المركب I والمركب III و IV المعقد لتدوير V المعقد ، وبالتالي توليد ATP9.
الأهم من ذلك ، أن نقل الإلكترون لا يحدث فقط بطريقة خطية ، ويشار إليه بخلاف ذلك باسم سلسلة نقل الإلكترون. بدلا من ذلك ، يمكن نقل الإلكترونات إلى تجمع الإنزيم المساعد Q من خلال مسارات تنفسية متعددة وتسهيل تدفق الإلكترونات المتقاربة. على سبيل المثال ، يمكن أن تدخل ركائز NADH و Uccinate عبر المركب I و II المعقد ، على التوالي. يمكن التبرع بالإلكترونات الناتجة عن أكسدة الأحماض الدهنية عبر مركب فلافوبروتين الناقل للإلكترون. والواقع أن التحليل الشامل ل OXPHOS يتطلب نهجا شاملا مع ركائز الوقود المناسبة (الشكل 1 ألف).
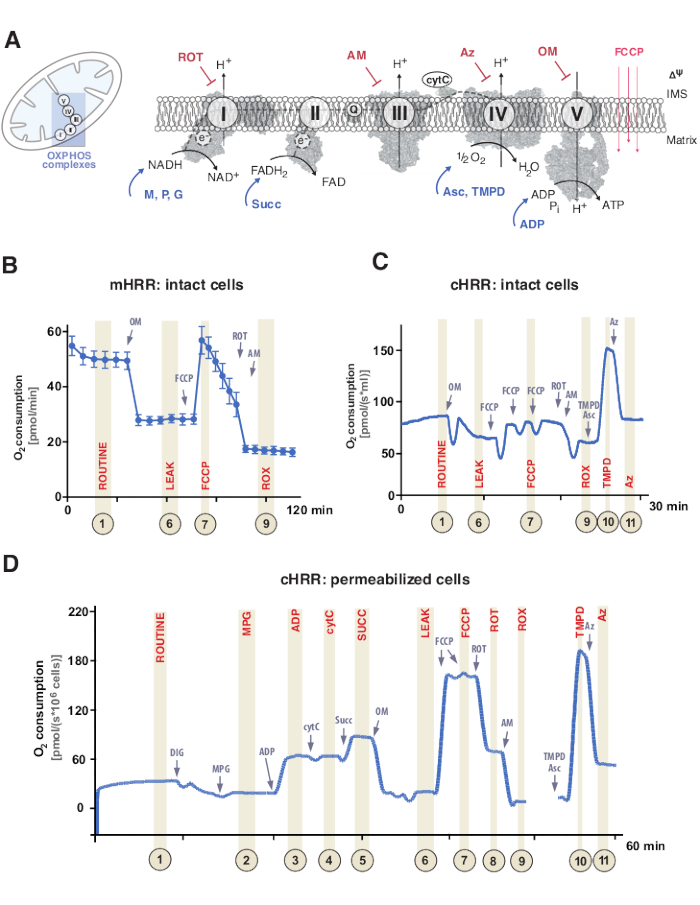
الشكل 1: الفسفرة التأكسدية للميتوكوندريا وبروتوكولات الركيزة والمثبطات المحددة. (أ) الميتوكوندريا ومخطط نظام نقل الإلكترون (CI-CIV) والميتوكوندريا F1F0 ATP synthase (CV). جميع الهياكل من PDB. الأرقام تصور فقط الركائز والمثبطات الموصوفة في هذه الدراسة). (ب) تتبع عينة من تدفق الأكسجين في خلايا HEK293 سليمة باستخدام بروتوكول قياسي في جهاز mHRR. (ج) تتبع عينة من تدفق الأكسجين في خلايا HEK293 سليمة باستخدام بروتوكول قياسي في جهاز cHRR. (د) تتبع عينة من تدفق الأكسجين في الخلايا الليفية البشرية المتخلل من متبرع سليم مع بروتوكول SUIT المعني. الاختصارات: 1 = التنفس الروتيني للخلايا السليمة. 2 = الدولة 2; 3 = الدولة 3 (طاء)؛ 4 = الحالة 3 (I) مع cytC ؛ 5 = الدولة 3 (I+II)؛ 6 = تسرب (OM); 7 = قدرة ETS; 8 = S (ROT) ؛ 9 = ROX; 10 = TMPD; 11 = Az. ROT = روتينون, AM = أنتيمايسين, ATP = أدينوسين ثلاثي الفوسفات, Az = أزيد, OM = أوليغومايسين, FCCP = سيانيد الكربونيل p-trifluoro-methoxyphenyl-hydrazone; Asc = أسكوربات، TMPD = N,N,N′,N′-رباعي ميثيل-P-فينيلينديامين، سوك = سكسينات، M = مالات، P = بيروفات، ADP = ثنائي فوسفات الأدينوزين، NAD = نيكوتيناميد الأدينين ثنائي النوكليوتيد، IMS = الفضاء بين الأغشية، FAD = فلافين الأدينين ثنائي النوكليوتيد. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
أصبح تحليل قدرة الميتوكوندريا OXPHOS باستخدام HRR طريقة كيميائية حيوية مفيدة ذات قيمة تشخيصية ليس فقط لعيوب الميتوكوندريا الأولية 10,11 ولكن تمتد إلى جميع مجالات البيولوجيا الأخرى مثل السرطان والشيخوخة12. يسمح HRR بتحديد التنفس الخلوي عن طريق تحليل قدرة OXPHOS للميتوكوندريا ، والتي تعكس بشكل مباشر نقص مجمع الجهاز التنفسي الفردي أو المشترك للميتوكوندريا ، ويرتبط بشكل غير مباشر بالخلل الوظيفي الخلوي وتغيير استقلاب الطاقة9. من الناحية المنهجية ، يتم إجراء قياسات التنفس باستخدام الخلايا أو الأنسجة أو الميتوكوندريا المعزولة11،13،14 ، مع المواد المجمدة المناسبة جزئيا فقط15،16. يظهر أن الأنسجة المجمدة لديها ETS سليمة مع الحفاظ على استقرار فائق التعقيد15. وبالتالي ، على عكس وسيطات TCA التقليدية ، يتم تغذية الركائز المعنية مباشرة في ETS. ومع ذلك ، يتم فقدان الاقتران بين ETS و ATP التوليف حيث يتم اختراق سلامة الغشاء من خلال تلف التجميد (تكوين بلورة الجليد).
عادة ما تتم تجارب التنفس عند درجة حرارة فسيولوجية تبلغ 37 درجة مئوية للحرارة الداخلية في الخلايا أو الأنسجة غير المتخلل أو المتغلغل. في حين أن الأول يأخذ في الاعتبار السياق الأيضي الخلوي ، فإن الأخير يوفر المساهمة النشطة لمجمعات OXPHOS الفردية و ATPase من خلال إضافة ركائز محددة (ومثبطات). أدى تسلسل وتباين الركائز والمثبطات إلى تطوير مجموعة متنوعة من بروتوكولات SUIT17 والمقايسات 18 لمعالجة مختلف المسائل العلمية لوظيفة OXPHOS (تمت مراجعتها تحت12). يقيم البروتوكول الأساسي للتنفس الخلوي أربع حالات مختلفة: أ) التنفس الروتيني – التنفس في وسائط التنفس المعنية دون أي إضافة للركائز أو المثبطات التي تستهلك ركائز داخلية المنشأ. يمكن أن تكشف هذه الحالة عن عيوب OXPHOS العامة أو عيوب التنفس الناجمة عن الثانوية ، على سبيل المثال ، بسبب تغيير ملامح الأيض. بعد ذلك ، تكشف إضافة مثبط ATPase oligomycin عن نفاذية غشاء الميتوكوندريا الداخلي للبروتونات ، والتي تعرف بأنها ii) تنفس التسرب. يسمح المعايرة اللاحقة للبروتونوفور مثل سيانيد الكربونيل غير المقترن p-trifluoro-methoxyphenyl-hydrazone (FCCP) بتحديد الحالة التي تكون فيها قدرة ETS قصوى في وضع دائرة البروتون المفتوحة عبر الغشاء ، والتي تعرف بأنها iii) التنفس غير المقترن. الأهم من ذلك ، يمكن أن تحدث حالة غير مقترنة أيضا عن طريق التدخلات التجريبية من خلال الأضرار الميكانيكية المفرطة لأغشية الميتوكوندريا. على العكس من ذلك ، تشير الحالة غير المقترنة إلى فك الارتباط التنفسي بواسطة آلية جوهرية يتم التحكم فيها من الناحية الفسيولوجية. وأخيرا، فإن التثبيط الكامل ل ETS بإضافة مضاد المايسين المثبط الثالث المعقد ومثبط الروتينون المعقد I يحدد استهلاك الأكسجين المتبقي (ROX) من العمليات غير المستهلكة للأكسجين الميتوكوندريا (الشكل 1A-C).
تتكون الطاقة الحيوية للميتوكوندريا من خمس حالات تنفس متميزة19,20. الحالة 1 التنفس بدون أي ركائز إضافية أو ADP ، باستثناء ما هو متاح داخليا. بعد إضافة ADP ، ولكن لا يزال ، لا توجد ركائز ، يتم تحقيق التنفس الحالة 2. عند إضافة الركائز ، مما يسمح بنقل الإلكترون وتخليق ATP ، يتم الوصول إلى التنفس في الحالة 3. في هذه الحالة ، يمكن تحديد قدرة OXPHOS عند التركيزات المشبعة من ADP ، والفوسفات غير العضوي ، والأكسجين ، والركائز المرتبطة ب NADH والسكسينات. يمكن تعريف التنفس في الحالة 4 أو التنفس LEAK على أنه حالة بدون ADP أو سينثاز ATP مثبط كيميائيا مع وجود ركائز كافية. أخيرا ، عندما يتم استنفاد كل الأكسجين (نقص الأكسجين) في إعداد غرفة مغلقة ، يلاحظ تنفس الحالة 5.
توجد عدة طرق لتقييم حالات الطاقة الخلوية14 مع جهازين يهيمنان على التقييم الحالي في الوقت الفعلي ل OXPHOS من خلال تحليل استهلاك الأكسجين ، ويتم قياسه كوظيفة لانخفاض الأكسجين بمرور الوقت في نظام غرفة مغلقة مع قابلية تطبيق مختلفة تعتمد على النموذج التجريبي وسؤال البحث: مقياس التنفس عالي الدقة Oroboros 2k ومحلل التدفق خارج الخلية Seahorse XF. يسجل كلا الجهازين معدلات استهلاك الأكسجين كانخفاض في البيكومولات (pmol) من الأكسجين (O2) في الثانية كقيمة مطلقة داخل الغرفة أو بئر microplate. يتم الحصول على استهلاك الأكسجين المحدد لكل كتلة عن طريق تطبيع استهلاك الأكسجين المعني في وصفة عازلة محددة لكل عدد من الخلايا (الملايين) أو وزن الأنسجة (ملغ) أو كمية البروتين.
O2k (Oroboros Instruments) هو نظام مغلق من غرفتين مجهز بمستشعر أكسجين بولوغرافي (يختصر باسم مقياس التنفس عالي الدقة القائم على الغرفة: cHRR). تحتوي كل غرفة تجريبية على 2 مل من السائل الذي يتم الاحتفاظ به متجانسا بواسطة أجهزة التقليب المغناطيسية. يستخدم مستشعر الأكسجين القطبي نهجا أمبيرومتري لقياس الأكسجين: فهو يحتوي على كاثود الذهب ، وأنود كلوريد الفضة / الفضة ، وبين محلول KCI الذي يخلق خلية كهروكيميائية يتم تطبيق الجهد عليها (0.8 فولت). ينتشر الأكسجين من وسط الفحص من خلال غشاء بروبيلين الإيثيلين المفلور 25 ميكرومتر (O 2-permeable) ويخضع للتخفيض عند الكاثود ، مما ينتج بيروكسيد الهيدروجين. في الأنود ، تتأكسد الفضة بواسطة بيروكسيد الهيدروجين ، مما يولد تيارا كهربائيا. يرتبط هذا التيار الكهربائي (الأمبير) خطيا بضغط الأكسجين الجزئي. يتم استخدام الضغط الجزئي للأكسجين وعامل ذوبان الأكسجين في وسط الفحص لحساب تركيز الأكسجين. نظرا لأن الضغط الجزئي للأكسجين يعتمد على درجة الحرارة التجريبية والقياسات المستقطبة حساسة لدرجة الحرارة ، فإن التقلبات في درجة الحرارة تحتاج إلى تنظيم دقيق (±0.002 درجة مئوية) بواسطة كتلة تسخين بلتييه. يمكن التحكم في درجة الحرارة ضمن نطاق 4 درجات مئوية و 47 درجة مئوية.
محلل التدفق خارج الخلية Seahorse XF (Agilent) هو نظام قائم على الألواح بتنسيق صفيحة دقيقة من 24 أو 96 بئرا حيث تقوم ثلاثة أقطاب كهربائية فلورية بقياس استهلاك الأكسجين بمرور الوقت في كل بئر (يختصر باسم مقياس التنفس عالي الدقة القائم على الصفائح الدقيقة: mHRR). يتوفر أربعة منافذ كحد أقصى في خرطوشة الفحص للحقن التلقائي أثناء الفحص. يحتوي الفحص على دورات متعددة ، لكل منها ثلاث مراحل: 1) الخلط ، 2) الانتظار ، و 3) القياس. خلال مرحلة القياس ، يتم خفض مجسات الاستشعار في الصفيحة الدقيقة مما يخلق غرفة مغلقة مؤقتا تحتوي على حجم 7-10 ميكرولتر لقياس الضوء المنبعث. ينبعث هذا الضوء من الفلوروفورات المدمجة في البوليمر على طرف مجسات المستشعر ، والتي تستشعر O2 بناء على تبريد الفوسفور. تتناسب شدة إشارة التألق مع O2 وتتأثر بدرجة حرارة المستشعر ووسط الفحص. لذلك ، يتطلب تقدير الأكسجين الدقيق نهجا نسبيا مع خلفية جيدة دون أي عينة. تحدث استعادة تركيز الأكسجين أثناء مرحلة الخلط عندما يتحرك المستشعر لأعلى ولأسفل لخلط الحجم فوق الغرفة المؤقتة. تحسب كل دورة معدل استهلاك أكسجين واحد. يمكن التحكم في درجة الحرارة ضمن نطاق 16 درجة مئوية و 42 درجة مئوية.
HRR هو المعيار الذهبي لتقييم الطاقة الحيوية الخلوية في الأمراض الأولية والمرتبطة بالميتوكوندريا والتمثيل الغذائي الخلوي العام. في هذه الدراسة ، يتم توفير البروتوكولات الأساسية ل HRR لتقييم وظيفة OXPHOS في الخلايا والأنسجة.
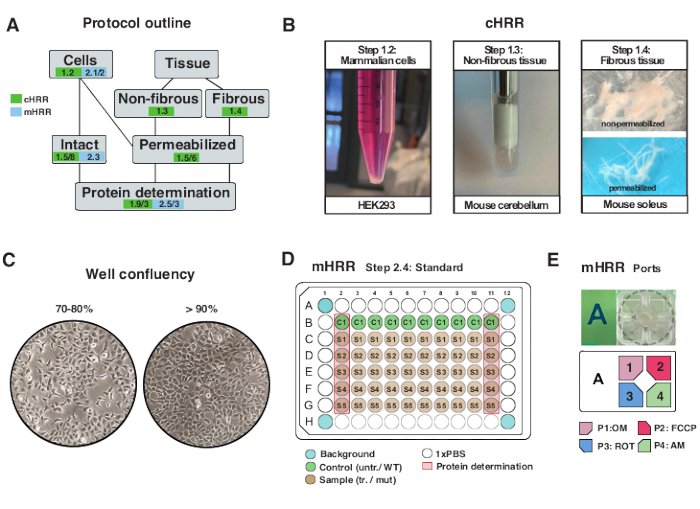
الشكل 2: سير العمل لتحضير الخلايا والأنسجة ل cHRR ، وإعداد الخلايا لقياس التنفس mHRR . (أ) مخطط البروتوكولات المقدمة. (ب) خلايا الثدييات (الخطوة 1.2): حبيبة HEK293 تساوي 3 × 106 خلايا (اللوحة اليسرى). الأنسجة غير الليفية (الخطوة 1.3): تحضير محللات المخيخ الفئرانية في 2 مل من بوتر تفلون (اللوحة الوسطى). نفاذية العضلات الهيكلية التي يسببها الصابونين (الخطوة 1.4) اللوحة اليمنى) لقياس التنفس cHRR. (ج) تخطيط بذر الصفائح الدقيقة القياسي (الخطوة 2.4) وفحص الالتقاء لتحليل الخلايا حقيقية النواة (HEK293) لقياس التنفس mHRR. (دال، هاء) مخطط تحميل منفذ الحقن لقياس التنفس mHRR (الخطوة 2.4). يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
