Mitocôndrias cumprem a oferta-chave de energia e são uma organela compartimentalizada contribuindo para processos bioenergetics celulares essenciais e metabólicos, como anabolismo de nucleotídeos, lipídios e aminoácidos, biogênese do aglomerado de enxofre de ferro e estão implicados em sinalização como morte celular controlada 1,2,3 . A bioenergeticia mitocondrial através da fosforilação oxidativa contribui para quase todos os processos celulares dentro da célula e, consequentemente, disfunções mitocondriais de origem primária ou secundária estão associadas a um amplo espectro de condições da doença 4,5. A disfunção mitocondrial envolve não apenas alterações na estrutura ou densidade mitocondrial, mas também na qualidade e regulação do sistema respiratório6. Este elemento qualitativo abrange controle de substrato, características de acoplamento, modificações pós-translacionais, dinâmica cristae e supercomplexos respiratórios 7,8. Portanto, a análise precisa da bioenergetics mitocondrial para abordagens experimentais e diagnósticas para avaliar o metabolismo energético da célula é importante na saúde e na doença.
Fosforilação oxidativa mitocondrial (OXPHOS) é uma sequência de reações dentro do sistema respiratório ou sistema de transferência de elétrons (ETS) para a geração de energia celular através de triptosfato de adenosina (ATP)9. O passo multienzimático para aproveitar a energia do fluxo de elétrons através dos complexos I e II ao complexo IV gera um gradiente eletroquímico de prótons através da membrana mitocondrial interna, posteriormente utilizado para fosforilação de difosfato de adenosina (ADP) para ATP via complexo V (F1FO ATP synthase) (Figura 1A).
Primeiro, os portadores de dois elétrons são gerados durante o ciclo tricarboxílico (TCA), glicolise e oxidação piruvato: dinucleotídeo de adenina de nicotinamida (NADH) e dinucleotídeo de adenina dihidroflavina (FADH2). O NADH é oxidado no complexo I (NADH desidrogenase), durante o qual dois elétrons são transferidos para a coenzima Q (quinona é reduzida a quinol), enquanto prótons são bombeados para o espaço intermembrano (IMS). Em segundo lugar, o complexo II (Succinato desidrogenase) oxida FADH2 e alimenta os elétrons para coenzima Q sem bombear prótons. Em terceiro lugar, no complexo III (Cytochrome c oxidoreductase), elétrons da coenzima Q são transferidos para citocromo c enquanto prótons são bombeados para o IMS. Em quarto lugar, o citocromo c transfere os elétrons para o complexo IV (Cytochrome c oxidase), o complexo final para bombear prótons, e onde o oxigênio funciona como um aceitador de elétrons para assimilar prótons, formando água. É esse oxigênio que as mitocôndrias consomem que pode ser medido por um oxígrafo. Finalmente, os prótons gerados a partir do complexo I, complexo III e complexo IV são usados para rodar o complexo V, gerando ASSIM ATP9.
É importante ressaltar que a transferência de elétrons ocorre não apenas de forma linear, de outra forma denotada como a cadeia de transporte de elétrons. Em vez disso, os elétrons podem ser transferidos para a piscina coenzima Q através de múltiplas vias respiratórias e facilitar o fluxo de elétrons convergentes. Os substratos e succinatos do NADH, por exemplo, podem entrar por meio do complexo I e do complexo II, respectivamente. Elétrons da oxidação de ácidos graxos podem ser doados através do complexo de flavoproteína de transferência de elétrons. De fato, uma análise abrangente do OXPHOS requer uma abordagem holística com substratos de combustível apropriados (Figura 1A).
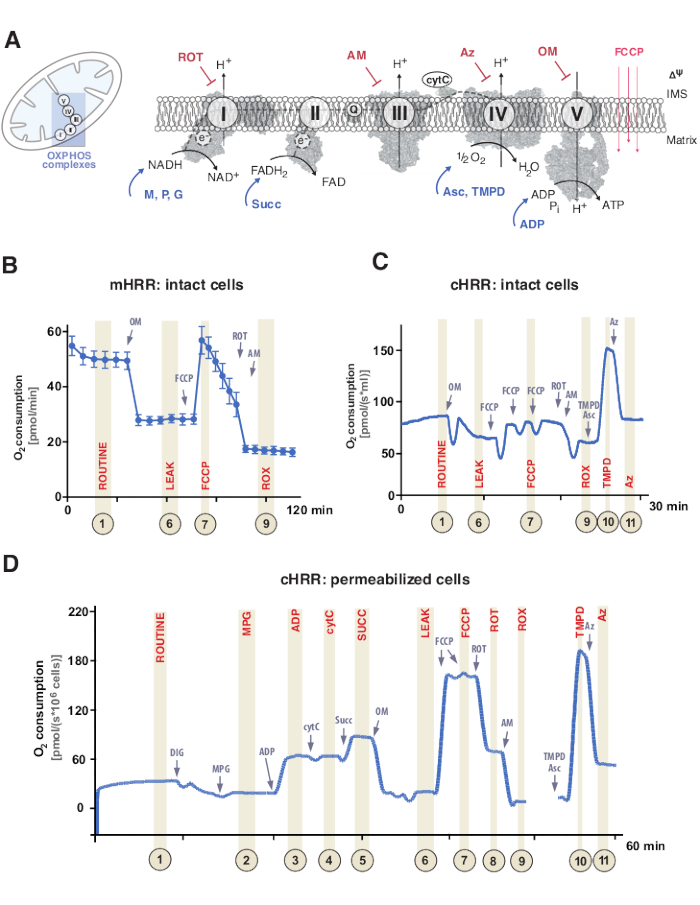
Figura 1: Fosforilação oxidativa mitocondrial e protocolos específicos de substrato e inibidor. (A) Mitoconddrion e esquema do sistema de transferência de elétrons (CI-CIV) e mitocondrial F1F0 ATP synthase (CV). Todas as estruturas são do PDB. As figuras retratam apenas substratos e inibidores descritos neste estudo). (B) Traços de amostra de fluxo de oxigênio em células HEK293 intactas usando protocolo padrão em um dispositivo mHRR. (C) Traços amostrais de fluxo de oxigênio em células HEK293 intactas usando protocolo padrão em um dispositivo cHRR. (D) Traços amostrais de fluxo de oxigênio em fibroblastos humanos permeabilizados de um doador saudável com o respectivo protocolo SUIT. Abreviaturas: 1 = Respiração rotineira de células intactas; 2 = Estado 2; 3 = Estado 3(I); 4 = Estado 3(I) com cítara; 5 = Estado 3 (I+II); 6 = Vazamento(OM); 7 = Capacidade de ETS; 8 = S(ROT); 9 = ROX; 10 = TMPD; 11 = Az. ROT = Rotenone, AM = Antimicina, ATP = Adenosina triphosfato, Az = Azide, OM = Oligomicina, FCCP = Cianeto carbonilizado p-trifluoro-metoxifenil-hidratante; Asc = Ascorbate, TMPD = N,N,N′,N’-tetramethyl-p-fenilelenediamina, Succ = Succinato, M = Malato, P = Piruado, ADP = Difosfato de Adenosina, NAD = Nicotinamida adenina dinucleotídeo, IMS = Espaço Intermembrano, FAD = Flavin adenina dinucleotídeo. Clique aqui para ver uma versão maior desta figura.
A análise da capacidade mitocondrial oxphos usando HRR tornou-se um método bioquímico instrumental de valor diagnóstico não apenas para defeitos mitocondriais primários10,11, mas estendendo-se a todos os outros domínios da biologia, como câncer e envelhecimento12. O HRR permite a determinação da respiração celular pela análise da capacidade mitocondrial de OXPHOS, que reflete diretamente deficiência do complexo respiratório mitocondrial individual ou combinado, e indiretamente está associada à disfunção celular e ao metabolismo energético alterado9. Metodologicamente, as medidas respiratórias são realizadas utilizando células, tecidos ou mitocôndrias isoladas 11,13,14, com material congelado apenas parcialmente adequado15,16. O tecido congelado é mostrado ter um ETS intacto com estabilidade de supercomplex mantido15. Assim, ao contrário dos intermediários tradicionais do TCA, os respectivos substratos são diretamente alimentados no ETS. No entanto, o acoplamento entre a síntese de ETS e ATP é perdido à medida que a integridade da membrana é comprometida por danos congelantes (formação de cristais de gelo).
Experimentos de respiração normalmente ocorrem a uma temperatura fisiológica de 37 °C para endotermias em células ou tecidos não permeabilizados ou permeabilizados. Enquanto o primeiro considera o contexto metabólico citosomático, este último fornece a contribuição energética dos complexos oxphos individuais e do ATPase através da adição de substratos específicos (e inibidores). A sequência e variação de substratos e inibidores levaram ao desenvolvimento de uma matriz diversificada de protocolos SUIT17 e ensaios18 para abordar várias questões científicas da função OXPHOS (revisadas abaixo de12). O protocolo básico de respiração celular avalia quatro estados diferentes: i) respiração de rotina – a respiração em uma respectiva mídia respiratória sem qualquer adição de substratos ou inibidores que consumam, mas substratos endógenos. Este estado pode revelar defeitos de respiração geralmente ou induzidos por secundários causados, por exemplo, por perfis metabólitos alterados. Em seguida, a adição do oligomicina inibidora de ATPase revela a permeabilidade da membrana mitocondrial interna aos prótons, definida como ii) respiração de vazamento. A titulação subsequente de um prótonophore como o desacontedor cianeto de cyanide carbonyl p-trifluoro-metoxiphenyl-hydrazone (FCCP) permite determinar o estado em que a capacidade de ETS é máxima em um modo de circuito de próton aberto transmembrano, definido como iii) respiração nãocopada. É importante ressaltar que um estado desacoplado também pode ocorrer por intervenções experimentais através de danos mecânicos excessivos às membranas mitocondriais. Por outro lado, o estado não acoplado refere-se ao desacoplamento respiratório por um mecanismo intrínseco que é fisiologicamente controlado. Finalmente, a inibição completa do ETS por adição do complexo inibidor III antimicina e do inibidor complexo I rotenona determina o consumo residual de oxigênio (ROX) de processos não mitocondriais de consumo de oxigênio (Figura 1A-C).
A bioenergetics mitocondrial consiste em cinco estados de respiração distintos19,20. A respiração do estado 1 é sem substratos adicionais ou ADP, exceto pelo que está disponível de forma endógena. Após a adição de ADP, mas ainda assim, sem substratos, a respiração estadual 2 é alcançada. Quando substratos são adicionados, permitindo transferência de elétrons e síntese ATP, a respiração do estado 3 é atingida. Neste estado, a capacidade oxphos pode ser definida em concentrações saturadas de ADP, fosfato inorgânico, oxigênio, nadh- e substratos ligados ao succinato. A respiração do estado 4 ou a respiração DE VAZAMENTO podem ser definidas como um estado sem ADP ou sintetizadores ATP quimicamente inibidos enquanto têm substratos suficientes. Por fim, quando todo o oxigênio é esgotado (anoxico) em um ambiente de câmara fechada, o estado 5 respiração é observado.
Existem vários métodos para avaliar os estados de energia celular14 com dois dispositivos dominando a avaliação atual em tempo real do OXPHOS através da análise do consumo de oxigênio, medido como função da diminuição do oxigênio ao longo do tempo em um sistema de câmara fechada com diferentes aplicabilidade dependentes do modelo experimental e da questão da pesquisa: o respirômetro oroboros de alta resolução e o analisador de fluxo extracelular Seahorse XF. Ambos os dispositivos registram as taxas de consumo de oxigênio como uma diminuição nos picomoles (pmol) de oxigênio (O2) por segundo como um valor absoluto dentro da câmara ou poço de microplato. O consumo específico de oxigênio por massa é obtido pela normalização do respectivo consumo de oxigênio em uma receita tampão específica por número de células (milhões), peso tecidual (mg) ou quantidade de proteína.
O O2k (Instrumentos Oroboros) é um sistema fechado de duas câmaras equipado com um sensor de oxigênio polarográfico (abreviado como respirômetro de alta resolução baseado em câmara: cHRR). Cada câmara experimental contém 2 mL de líquido que é mantido homogêneo por agitadores magnéticos. O sensor de oxigênio polarográfico utiliza uma abordagem amperométrica para medir o oxigênio: contém um cátodo de ouro, um ânodo de cloreto de prata/prata, e entre uma solução KCI criando uma célula eletroquímica sobre a qual uma tensão (0,8 V) é aplicada. O oxigênio do ensaio difere através de uma membrana fluorina de etileno fluorado de 25 μm (O 2-permeável) e sofre redução no cátodo, produzindo peróxido de hidrogênio. No ânodo, a prata é oxidada por peróxido de hidrogênio, gerando uma corrente elétrica. Esta corrente elétrica (ampere) está linearmente relacionada com a pressão parcial de oxigênio. A pressão parcial do oxigênio e o fator de solubilidade de oxigênio do meio de ensaio são usados para calcular a concentração de oxigênio. Uma vez que a pressão parcial do oxigênio depende da temperatura experimental e as medidas polarográficas são sensíveis à temperatura, as flutuações na temperatura precisam de regulação precisa (±0,002 °C) por um bloco de aquecimento Peltier. A temperatura pode ser controlada dentro de uma faixa de 4 °C e 47 °C.
O analisador de fluxo extracelular Seahorse XF (Agilent) é um sistema baseado em placas com formato de microplaca de 24 ou 96 poços no qual três eletrodos de fluorescência medem o consumo de oxigênio ao longo do tempo em cada poço (abreviado como respirômetro de alta resolução baseado em microplaca: mHRR). Um máximo de quatro portas no cartucho de ensaio estão disponíveis para injeção automatizada durante o ensaio. Um ensaio contém vários ciclos, cada um com três fases: 1) mistura, 2) espera e 3) medição. Durante a fase de medição, as sondas de sensores são reduzidas na microplacão criando uma câmara temporariamente fechada contendo volume de 7-10 μL para medir a luz emitida. Esta luz é emitida por fluoroforos embutidos em polímero na ponta das sondas do sensor, que sentem O2 com base na saciamento da fosforescência. A intensidade do sinal de fluorescência é proporcional a O2 e influenciada pela temperatura do sensor e do meio de ensaio. Portanto, a estimativa precisa de oxigênio requer uma abordagem relativa com um poço de fundo sem qualquer amostra. A restauração da concentração de oxigênio ocorre durante a fase de mistura quando o sensor se move para cima e para baixo para misturar o volume acima da câmara temporária. Cada ciclo calcula uma taxa de consumo de oxigênio. A temperatura pode ser controlada dentro de uma faixa de 16 °C e 42 °C.
O HRR é o padrão-ouro para avaliar bioenergetics celulares em doenças primárias e mitocôndrias associadas e metabolismo celular geral. Neste estudo, protocolos básicos para HRR são fornecidos para avaliar a função OXPHOS em células e tecidos.
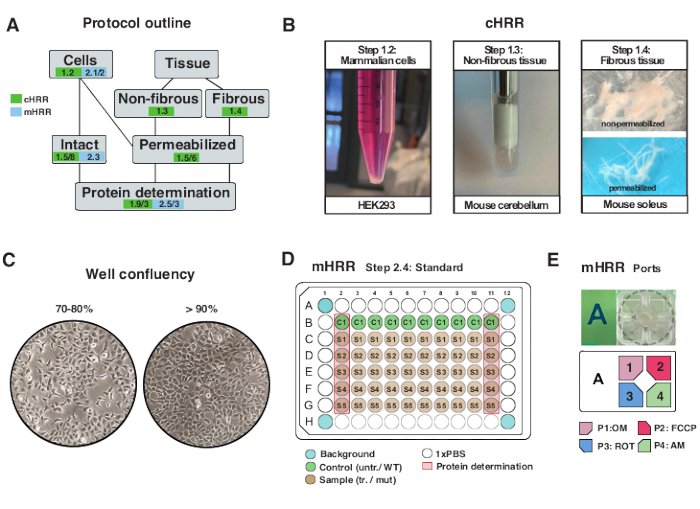
Figura 2: Fluxo de trabalho para preparações celulares e tecidos para rCR, e preparação celular para respirometria mHRR. (A) Esboço dos protocolos fornecidos. (B) Células mamíferas (passo 1.2): pelota HEK293 igual a 3 x 106 células (painel esquerdo). Tecido não fibroso (passo 1.3): Preparação de cerebelo murino em 2 mL Teflon potter (painel médio). Permeabilização do músculo esquelético induzida por sapona (etapa 1.4) painel direito) para respirometria de HRR. (C) Layout de semeamento de microplaca padrão (etapa 2.4) e verificação de confluência para a análise de células eucarióticas (HEK293) para respirometria mHRR. (D, E) Esquema de carregamento da porta de injeção para respirometria mHRR (etapa 2.4). Clique aqui para ver uma versão maior desta figura.
