Investigating the Protective Effects of Platycodin D on Non-Alcoholic Fatty Liver Disease in a Palmitic Acid-Induced In Vitro Model
Summary
This protocol investigates the protective effects of platycodin D on non-alcoholic fatty liver disease in a palmitic acid-induced in vitro model.
Abstract
The occurrence of non-alcoholic fatty liver disease (NAFLD) has been increasing at an alarming rate worldwide. Platycodon grandiflorum is widely used as a traditional ethnomedicine for the treatment of various diseases and is a typical functional food that can be incorporated into the everyday diet. Studies have suggested that platycodin D (PD), one of the main active ingredients in Platycodon grandiflorum, has high bioavailability and significantly mitigates the progress of NAFLD, but the underlying mechanism of this is still unclear. This study aims to investigate the therapeutic effect of PD against NAFLD in vitro. AML-12 cells were pretreated with 300 µM palmitic acid (PA) for 24 h to model NAFLD in vitro. Then, the cells were either treated with PD or received no PD treatment for 24 h. The levels of reactive oxygen species (ROS) were analyzed using 2′,7′-dichloro-dihydro-fluorescein diacetate (DCFH-DA) staining, and the mitochondrial membrane potential was determined by the JC-1 staining method. Moreover, the protein expression levels of LC3-II/LC3-I and p62/SQSTM1 in the cell lysates were analyzed by western blotting. PD was found to significantly decrease the ROS and mitochondrial membrane potential levels in the PA-treated group compared to the control group. Meanwhile, PD increased the LC3-II/LC3-I levels and decreased the p62/SQSTM1 levels in the PA-treated group compared to the control group. The results indicated that PD ameliorated NAFLD in vitro by reducing oxidative stress and stimulating autophagy. This in vitro model is a useful tool for studying the role of PD in NAFLD.
Introduction
Platycodon grandiflorus (PG), which is the dried root of Platycodon grandiflorus (Jacq.) A.DC., is used in traditional Chinese medicine (TCM). It is mainly produced in the northeast, north, east, central, and southwest regions of China1. The PG components include triterpenoid saponins, polysaccharides, flavonoids, polyphenols, polyethylene glycols, volatile oils, and minerals2. PG has a long history of being used as a food and a herbal medicine in Asia. Traditionally, this herb was used to make medicine against lung diseases. Modern pharmacology also provides evidence of the efficacy of PG for treating other diseases. Studies have shown that PG has a therapeutic effect on a variety of drug-induced liver injury models. The dietary supplementation of PG or platycodin extracts can ameliorate high-fat diet-induced obesity and its related metabolic diseases3,4,5. Polysaccharides from PG can be used for the treatment of acute liver injury caused by LPS/D-GalN in mice6. Moreover, saponins from the roots of PG ameliorate high-fat diet-induced non-alcoholic steatohepatitis (NASH)7. Furthermore, platycodin D (PD), one of the most important therapeutic components of PG, can enhance low-density lipoprotein receptor expression and low-density lipoprotein uptake in human hepatocellular carcinoma (HepG2) cells8. Furthermore, PD can also induce apoptosis and inhibit adhesion, migration, and invasion in HepG2 cells9,10. Thus, in this study, mouse hepatoma AML-12 cells are used for in vitro model construction and to further study the pharmacological effects and underlying mechanisms of PD in this model.
The term non-alcoholic fatty liver disease (NAFLD) refers to a group of liver diseases that includes simple steatosis, NASH, cirrhosis, and hepatocellular carcinoma11. Although the pathogenesis of NAFLD is incompletely understood, from the classic "two-hit" theory to the current "multiple-hit" theory, insulin resistance is considered to be central in the pathogenesis of NAFLD12,13,14. Studies have demonstrated that insulin resistance in hepatocytes could lead to increased free fatty acids, which form triglycerides that are deposited in the liver and cause the liver to become fatty15,16. The accumulation of fat can lead to lipotoxicity, oxidative stress-induced mitochondrial dysfunction, endoplasmic reticulum stress, and inflammatory cytokine release, resulting in the pathogenesis and progression of NAFLD17,18. In addition, autophagy also plays a role in the pathogenesis of NAFLD, as it is involved in regulating cellular insulin sensitivity, cellular lipid metabolism, hepatocyte injury, and innate immunity19,20,21.
A variety of animal models and cellular models have been established to provide a basis for exploring the pathogenesis and potential therapeutic targets of NAFLD22,23. However, single animal models cannot fully mimic all the pathological processes of NAFLD24. Individual differences between animals lead to different pathological features. Using liver cell lines or primary hepatocytes in in vitro studies of NAFLD ensures maximum consistency in the experimental conditions. Hepatic lipid metabolism dysregulation can lead to higher levels of hepatocyte lipid droplet accumulation in NAFLD25. Free fatty acids such as oleic acid and palm oil have been used in the in vitro model to mimic NAFLD caused by a high-fat diet26,27. The human hepatoblastoma cell line HepG2 is often used in the construction of NAFLD models in vitro, but, as a tumor cell line, the metabolism of HepG2 cells is significantly different from that of liver cells under normal physiological conditions28. Therefore, using primary hepatocytes or mouse primary hepatocytes to construct the in vitro NAFLD model for drug screening is more advantageous than using tumor cell lines. Comparing the synergistic examination of drug effects and therapeutic targets in both animal models and in vitro hepatocyte models, it seems that using mouse hepatocytes to construct the in vitro NAFLD model has better application potential.
Free fatty acids entering the liver are oxidized to produce energy or stored as triglycerides. Significantly, free fatty acids have a certain lipotoxicity and may induce cellular dysfunction and apoptosis12. Palmitic acid (PA) is the most abundant saturated fatty acid in human plasma29. When cells in non-adipose tissue are exposed to high concentrations of PA for a long time, this stimulates the production of reactive oxygen species (ROS) and causes oxidative stress, lipid accumulation, and even apoptosis30. Therefore, many researchers use PA as an inducer to stimulate liver cells to produce ROS and, thus, construct the in vitro fatty liver disease model and evaluate the protective effects of certain active substances on cells31,32,33,34. This study introduces a protocol for investigating the protective effects of PD on a cell model of NAFLD induced by PA.
Protocol
AML-12 cells (a normal mouse hepatocyte cell line) are used for the cell-based studies. The cells are obtained from a commercial source (see Table of Materials).
1. Pretreatment of the AML-12 cells to model NAFLD in vitro
- Maintain the cells in normal cell culture media (DMEM plus Ham's F12 [1:1] containing 0.005 mg/mL insulin, 5 ng/mL selenium, 0.005 mg/mL transferrin, 40 ng/mL dexamethasone, and 10% fetal bovine serum [FBS], see Table of Materials) at 37 °C in a humidified atmosphere with 5% CO2.
- Seed the cells in 12-well plates (1 mL/well) with a density of 2.8 x 105 cells/well.
NOTE: All the cell digestion and medium exchange operations are performed in a biosafety cabinet to avoid contaminating the cells. - Remove the culture medium of the cells after the overnight incubation (~10 h), and then wash the cells two times with 1 mL of serum-free media (per well).
- Divide the 12-well cell plate into four different treatment groups from left to right, including (1) the control group, (2) the PD-treated group, (3) the PA-treated group, and (4) the PA + PD-treated group.
NOTE: Three replicates of each experimental treatment group are arranged from top to bottom on the same 12-well cell plate. - Add the normal cell culture media (1 mL/well) to the control group and the PD-treated group, and add the normal cell culture media (1 mL/well) supplemented with 300 µM of PA to the PA-treated and PA + PD-treated group.
- Remove the culture medium of the cells after 24 h of incubation, and then wash the cells with 1 mL of serum-free media (per well) two times.
- Add normal cell culture media (1 mL/well) supplemented with a vehicle (dimethyl sulfoxide, DMSO, 0.1% v/v) to the control group; add normal cell culture media (1 mL/well) supplemented with 1 µM PD to the PD-treated group; add normal cell culture media (1 mL/well) supplemented with 300 µM PA to the PA-treated group; add normal cell culture media (1 mL/well) supplemented with 300 µM PA and 1 µM PD to the PA + PD-treated group.
- After incubation for an additional 24 h, investigate the protective effects of PD on cells using 2′,7′-dichloro-dihydro-fluorescein diacetate (DCFH-DA) staining (step 2), JC-1 staining (step 3), and western blotting (step 4).
NOTE: All the incubation operations are done at 37 °C in a humidified atmosphere with 5% CO2.
2. Measurement of the change in ROS production
NOTE: The intracellular ROS levels in the cells are assessed based on the DCFH-DA staining assay.
- At the end of the treatment period (step 1.8), wash the cells with 1 mL of phosphate-buffered saline per well (1x PBS, pH 7.4) three times, and then stain the cells with 100 µL of 10 µM DCFH-DA per well (see Table of Materials). Incubate the cells in the dark for 30 min.
NOTE: If no obvious green fluorescence is observed after 30 min of incubation, the incubation time of the probe and cells can be appropriately increased (30-60 min). If overexposure of the green fluorescence value is observed after 30 min of incubation, the incubation time of the probe and cells can be appropriately reduced (15-30 min). - Wash the cells with 1x PBS (1 mL/well) three times. Add 1 mL of 1x PBS to each well.
- Place the 12-well plate on the microscope stage (see Table of Materials), and then use a 20x objective to observe the morphology of the cells (magnification: 200x).
- Capture three representative fluorescence images (magnification: 200x) for each well on the fluorescence microscope using the green fluorescent channel with an excitation wavelength of 480 nm and an emission wavelength of 530 nm.
NOTE: The green fluorescent channel with an excitation wavelength of 480 nm and an emission wavelength of 530 nm is recommended to detect the DCFH-DA. Additionally, DCFH-DA can be detected with the parameter settings for GFP and FITC in the fluorescence microscope35,36. - Finally, process the images with image-acquisition software (see Table of Materials), and then use ImageJ software to calculate the mean fluorescence intensity of each group or the ratios of the different groups.
NOTE: The technical details of the fluorescence microscope and Image J software used for the imaging work have been described previously37,38.
3. Measurement of the change in mitochondrial membrane potential
NOTE: The changes in mitochondrial membrane potential are monitored by the JC-1 staining assay.
- At the end of the treatment period (step 1.8), wash the cells with 1 mL of 1x PBS per well three times, and then stain the cells with 100 µL of 5 µg/mL JC-1 working solution (see Table of Materials) for 30 min at 37 °C in the dark.
NOTE: If no obvious green fluorescence is observed after 30 min of incubation, the incubation time of the probe and cells can be appropriately increased (30-60 min). If overexposure of the green fluorescence value is observed after 30 min of incubation, the incubation time of the probe and cells can be appropriately reduced (15-30 min). - Wash the cells with 1x PBS (1 mL/well) three times. Add 1 mL of 1x PBS to each well.
- Place the 12-well plate on the microscope stage, and then use a 20x objective to observe the morphology of the cells (magnification: 200x).
- Capture three representative fluorescence images (magnification: 200x) for each well on the fluorescence microscope using the green fluorescent channel with excitation and emission wavelengths of 485 nm and 535 nm, respectively, and the red fluorescent channel with excitation and emission wavelengths of 550 nm and 600 nm, respectively.
NOTE: The green fluorescent channel with an excitation wavelength of 485 nm and an emission wavelength of 535 nm is used to detect the JC-1 monomer, which is treated as depolarized mitochondria39,40,41, and the red fluorescent channel with an excitation wavelength of 550 nm and an emission wavelength of 600 nm is used to detect the JC-1 dimer, which is treated as polarized mitochondria39,40,41. - Finally, process the images with image-acquisition software, and then use Image J software to calculate the mean fluorescence intensity of each group or the ratios of the different groups.
NOTE: The technical details of the fluorescence microscope and Image J software used for the imaging work have been described previously37,38.
4. Measurement of the protein expression levels of LC3-II/LC3-I and p62/SQSTM1
- After treatment (step 1.8), wash the cells three times with 1x PBS (1 mL/well) that has been precooled at 4 °C.
- Add RIPA lysis buffer (100 µL/well) supplemented with protease and a phosphatase inhibitor cocktail (1x) (see Table of Materials) to the 12-well plate, and lyse on ice for 5 min.
- Collect the cell lysate into 1.5 mL microcentrifuge tubes, and centrifuge at 12,000 x g for 20 min at 4 °C. Determine the protein concentration of the supernatants by the BCA method following the standard procedures42.
- Add SDS-PAGE sample loading buffer (5x, see Table of Materials) to the cell lysate supernatants (volume ratio = 1:4).
- Mix by vortexing (at high speed for ~15 s), and heat the mixed samples at 100 °C for 5 min to denature the protein.
- Put the 12-well-prepared 12% SDS-PAGE gel into the electrophoresis tank, and then add the SDS up-sample buffer (1x) diluted with ultra-pure water to the height limit position.
NOTE: The SDS-PAGE gel is prepared using a commercial kit (see Table of Materials) according to the manufacturer's instructions. - Add the protein marker (5 µL/well) and samples (20 µg/well) into different wells of the SDS-PAGE gel.
- Set the stable voltage mode at 100 V, and perform the electrophoresis for 80 min.
- After the SDS-PAGE electrophoresis, carry out the electrotransfer of the proteins to a polyvinylidene fluoride (PVDF) membrane (0.45 µM, see Table of Materials) following previously published reports43,44.
- After the electrotransfer of the proteins, wash the PVDF membrane with 10 mL of TBST (1x TBS, 0.1% Tween 20) two times (5 min/time) in a shaker at room temperature.
- Block the PVDF membrane with 5 mL of bovine serum albumin (BSA, 5%) in a shaker at room temperature for 1 h.
- Wash the PVDF membrane with 10 mL of TBST three times (10 min/time). Then, incubate the PVDF membrane in 5 mL of the specific primary antibodies diluted in blocking buffer for LC3 (mouse mAb, 1:2,000), p62/SQSTM1 (hereafter referred to as p62, mouse mAb, 1:2,000), and β-actin (mouse mAb, 1:2,000) (see Table of Materials) overnight at 4 °C.
- Wash the PVDF membrane with 10 mL of TBST three times (10 min/time) at room temperature. Then, incubate the PVDF membrane with rabbit anti-mouse IgG (HRP) secondary antibody diluted in blocking buffer (1:10,000) (see Table of Materials) at room temperature and sheltered from light for 2 h.
- Wash the PVDF membrane with 10 mL of TBST three times (10 min/time) at room temperature. Then, place the PVDF membrane on the plastic wrap, add an appropriate amount of ECL working solution (200 µL/membrane) (see Table of Materials), and incubate for 2 min.
- After incubation, remove the ECL working solution, and expose the PVDF membrane in the imaging system for image development. Finally, use the Image J software to analyze the gray values of each band.
NOTE: The technical details of the western blotting and Image J software used for the imaging work have been described previously45,46.
5. Statistical analysis
- Present the data from the experiments as the mean ± standard deviation (SD).
- Perform the analysis of significance with the statistical software tool described previously47.
- Calculate the statistical differences between the two groups using a t-test. A P-value below 0.05 is considered statistically significant: *P < 0.05, **P < 0.01, ***P < 0.005.
Representative Results
Intracellular ROS in the cells
AML-12 cells were induced with 300 µM PA for 24 h, and a NAFLD cell model was established. Subsequently, the cells were treated with PD for 24 h. The cells were labeled with a DCFH-DA fluorescent probe, and ROS production was observed under a fluorescence microscope. The results of the DCFH-DA staining of intracellular ROS in the cells are shown in Figure 1. The results showed that PD could significantly reduce the level of intracellular ROS in the cells incubated with 300 µM of PA (P < 0.01), indicating that PD can reduce cellular oxidative stress.
Mitochondrial membrane potential of the cells
The mitochondrion is the central organelle governed by the intrinsic apoptosis pathway48,49,50. Therefore, the mitochondrial membrane potential (MMP) of AML-12 cells that had been incubated with PD for 24 h was observed under a fluorescence microscope after JC-1 staining. As shown in Figure 2, the green fluorescence (JC-1 monomers, treated as depolarized mitochondria39,40,41) in the PA-treated cells was higher than in the untreated cells. On the other hand, the red fluorescence (JC-1 dimers, treated as polarized mitochondria39,40,41) in the PA-treated cells was lower than in the untreated cells, indicating PA-induced MMP depolarization in the cells. In addition, compared with the model group, the ratio of JC-1 monomers:dimers decreased in the PD treatment group (P < 0.01), indicating that PD could ameliorate PA-induced MMP depolarization.
The protein expression levels of LC3-II/LC3-I and p62
This study also investigated the potential role of PD on the autophagy pathway by investigating the protein expression levels of autophagy-related proteins LC3 and p62 in the cells by western blotting. As shown in Figure 3, the ratio of LC3-II:LC3-I significantly decreased (P < 0.005), and the protein expression level of p62 increased after being treated with PA for 48 h (P < 0.01). On the other hand, PD could significantly reduce the protein expression level of p62 (P < 0.01) and increase the ratio of LC3-II:LC3-I (P < 0.05), indicating that PD may restore the autophagic flux inhibited by PA.
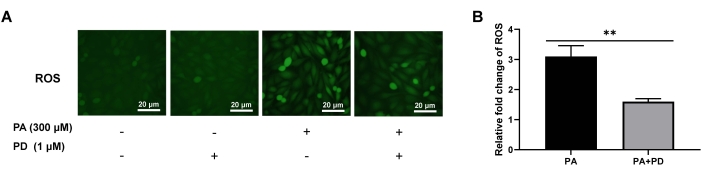
Figure 1: Effects of PD on intracellular ROS in the cells as assessed by DCFH-DA staining. (A) ROS generation was detected by fluorescence microscopy, 200x. Scale bar: 20 µm. (B) Relative fold changes in the ROS levels in different groups. Each column represents mean ± SD (n = 3). **P < 0.01. Please click here to view a larger version of this figure.
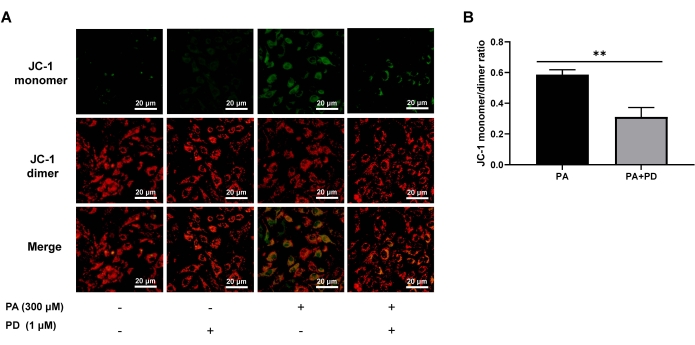
Figure 2: Effects of PD on MMP depolarization in the cells as assessed by JC-1 staining. (A) MMP depolarization was detected by fluorescence microscopy, 200x. Scale bar: 20 µm. (B) Relative changes in the JC-1 monomers:dimers ratio in the different groups. Each column represents mean ± SD (n = 3). **P < 0.01. Please click here to view a larger version of this figure.
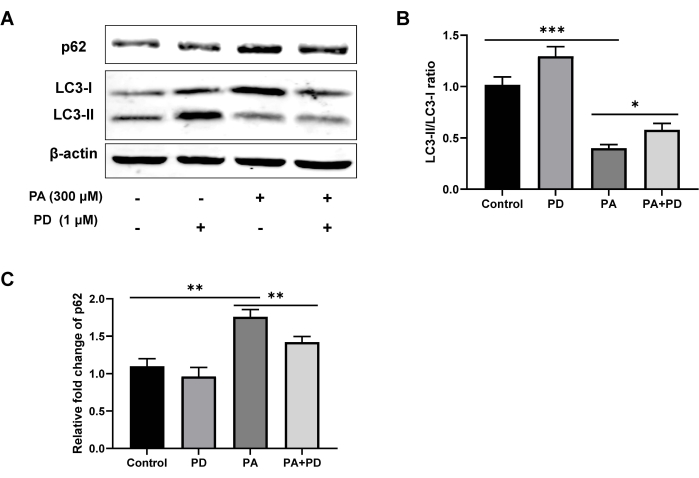
Figure 3: Effects of PD on the protein expression levels of LC3-II/LC3-I and p62 in the cells as assessed by western blotting. (A) The protein levels of LC3, p62, and β-actin were detected by western blotting. (B) Relative changes in the LC3-II:LC3-I ratio in the different groups. (C) Relative fold changes in the expression of p62 in the different groups. Each column represents mean ± SD (n = 3). *P < 0.05, **P < 0.01, ***P < 0.001. Please click here to view a larger version of this figure.
Discussion
Studies have highlighted the fact that NAFLD is a clinicopathological syndrome, ranging from fatty liver to NASH, that can progress to cirrhosis and liver cancer51. A high-fat diet and an inactive lifestyle are typical risk factors for NAFLD. Both non-drug therapies and drug therapies for NAFLD treatment have been researched51,52,53. However, the pathogenesis of NAFLD has not been fully elucidated. The method described here involves an in vitro model of NAFLD established by PA-stimulated AML-12 cells. We investigated the protective effects of PD against NAFLD in this model, and we found that PD attenuated the PA-induced increase in ROS. Besides, PD also ameliorated PA-induced MMP depolarization. Furthermore, PD could significantly reduce the protein expression level of p62 and increase the ratio of LC3-II:LC3-I. These experimental results could provide a basis for the application of PD as an active pharmaceutical ingredient for NAFLD treatment.
As a major intracellular degradation pathway, autophagy realizes the physiological process of recycling and reusing nutrient raw materials by degrading excess components in cells19,54. Autophagy is also a key repair mechanism for the body to maintain "yin-yang balance", and it is the microscopic embodiment of "qi transformation" in TCM55,56,57. In the present protocol, in addition to detecting the changes in ROS production and MMP depolarization, the potential role of PD on the autophagy pathway was investigated by detecting the protein expression levels of autophagy-related proteins LC-3 and p62 by western blotting. After the treatment of PA-induced cells with PD, the ratio of LC3-II:LC3-I decreased, and the accumulation of p62 increased, indicating that the level of autophagy decreased. The experimental results are consistent with other literature reports20,34,58,59. This in vitro cell model may help improve the understanding of the biological function of PD and help study the use of other active TCM substances for the treatment of NAFLD.
There are some critical steps in the experimental process. To determine whether the in vitro NAFLD model is successfully established, the lipid droplet deposits can be compared in the normal control and PA-treated group cells by Oil Red O staining, as described previously60,61. After DCFH-DA staining or JC-1 staining, the remaining probes that do not enter the cells must be washed off; otherwise, this will cause a high background when taking the fluorescence images. In addition, the time (except for incubation time) between the loading and measurement of the fluorescent probes (DCFH-DA or JC-1) must be shortened to reduce the possibility of experimental errors. Moreover, the working concentrations of the fluorescent probes (DCFH-DA or JC-1) can be adjusted as needed if the fluorescence value of the cells in the negative control group is relatively high. Alternatively, the incubation time of the JC-1 working solution and the cells can be appropriately adjusted within a timeframe of 15-60 min according to the actual experimental conditions.
There are also some limitations in this in vitro cell model. Cells such as hepatic parenchymal cells, hepatic stellate cells, Kupffer cells, and vascular endothelial cells are present in liver tissues62. Therefore, experiments only on hepatic parenchymal cells may not be adequate to explain the effects of drugs. Other cell models also need to be used in anti-NAFLD drug discovery. In addition, three-dimensional (3D) cell culture systems have been used to construct liver models and test hepatotoxicity63,64. Future studies will focus on developing in vitro models using co-culture and 3D cell culture techniques for potential anti-NAFLD drug screening.
As a multi-system disease, many factors play a role in the development and progression of NAFLD, meaning a single animal model or cell model cannot fully mimic all the pathological processes of this disease65. Over-reliance on the use of animal models increases the burden of therapeutic drug development. Using in vitro cell models in the early stages of anti-NAFLD drug development is more practical and cost-effective. In this study, a normal mouse hepatocyte cell line AML-12 was used to construct the in vitro model of NAFLD, and the therapeutic effect of PD was validated in this model. Notably, the results of this study provide a basis for further research on the anti-NAFLD effects of PD in mice hepatocyte and primary hepatocyte models.
In conclusion, an in vitro model to study the protective effects of PD against NAFLD is presented here. This also could be a useful in vitro model for investigating the efficacy of other active substances of TCM for the treatment of NAFLD.
Disclosures
The authors have nothing to disclose.
Acknowledgements
This work is supported by grants from the Chongqing Science and Technology Commission (cstc2020jxjl-jbky10002, jbky20200026, cstc2021jscx-dxwtBX0013, and jbky20210029) and the China Postdoctoral Science Foundation (No. 2021MD703919).
Materials
| 5% BSA Blocking Buffer | Solarbio, Beijing, China | SW3015 | |
| AML12 (alpha mouse liver 12) cell line | Procell Life Science&Technology Co., Ltd, China | AML12 | |
| Beyo ECL Plus | Beyotime, Shanghai, China | P0018S | |
| Bio-safety cabinet | Esco Micro Pte Ltd, Singapore | AC2-5S1 A2 | |
| cellSens | Olympus, Tokyo, Japan | 1.8 | |
| Culture CO2 Incubator | Esco Micro Pte Ltd, Singapore | CCL-170B-8 | |
| Dexamethasone | Beyotime, Shanghai, China | ST125 | |
| Dimethyl sulfoxide | Solarbio, Beijing, China | D8371 | |
| DMEM/F12 | Hyclone, Logan, UT, USA | SH30023.01 | |
| Foetal Bovine Serum | Hyclone, Tauranga, New Zealand | SH30406.05 | |
| Graphpad software | GraphPad Software Inc., San Diego, CA, USA | 8.0 | |
| HRP Goat Anti-Mouse IgG (H+L) | ABclonal, Wuhan, China | AS003 | |
| Hydrophobic PVDF Transfer Membrane | Merck, Darmstadt, Germany | IPFL00010 | |
| Insulin, Transferrin, Selenium Solution, 100× | Beyotime, Shanghai, China | C0341 | |
| MAP LC3β Antibody | Santa Cruz Biotechnology (Shanghai) Co., Ltd | SC-376404 | |
| Mitochondrial Membrane Potential Assay Kit with JC-1 | Solarbio, Beijing, China | M8650 | |
| Olympus Inverted Microscope IX53 | Olympus, Tokyo, Japan | IX53 | |
| Palmitic Acid | Sigma, Germany | P0500 | |
| Penicillin-Streptomycin Solution (100x) | Hyclone, Logan, UT, USA | SV30010 | |
| Phenylmethanesulfonyl fluoride | Beyotime, Shanghai, China | ST506 | |
| Phosphate Buffered Solution | Hyclone, Logan, UT, USA | BL302A | |
| Platycodin D | Chengdu Must Bio-Technology Co., Ltd, China | CSA: 58479-68-8 | |
| Protease inhibitor cocktail for general use, 100x | Beyotime, Shanghai, China | P1005 | |
| Protein Marker | Solarbio, Beijing, China | PR1910 | |
| Reactive Oxygen Species Assay Kit | Solarbio, Beijing, China | CA1410 | |
| RIPA Lysis Buffer | Beyotime, Shanghai, China | P0013E | |
| SDS-PAGE Gel Quick Preparation Kit | Beyotime, Shanghai, China | P0012AC | |
| SDS-PAGE Sample Loading Buffer, 5x | Beyotime, Shanghai, China | P0015 | |
| Sigma Centrifuge | Sigma, Germany | 3K15 | |
| SQSTM1/p62 Antibody | Santa Cruz Biotechnology (Shanghai) Co., Ltd | SC-28359 | |
| Tecan Infinite 200 PRO | Tecan Austria GmbH, Austria | 1510002987 | |
| WB Transfer Buffer,10x | Solarbio, Beijing, China | D1060 | |
| β-Actin Mouse mAb | ABclonal, Wuhan, China | AC004 |
References
- Xunyan, X. Y., Fang, X. M. The effect of Platycodon grandiflorum and its historical change in the clinical application of Platycodonis radix. Zhonghua Yi Shi Za Shi. 51 (3), 167-176 (2021).
- Ma, X., et al. Platycodon grandiflorum extract: Chemical composition and whitening, antioxidant, and anti-inflammatory effects. RSC Advances. 11 (18), 10814-10826 (2021).
- Ke, W., et al. Dietary Platycodon grandiflorus attenuates hepatic insulin resistance and oxidative stress in high-fat-diet induced non-alcoholic fatty liver disease. Nutrients. 12 (2), 480 (2020).
- Kim, Y. J., et al. Platycodon grandiflorus root extract attenuates body fat mass, hepatic steatosis and insulin resistance through the interplay between the liver and adipose tissue. Nutrients. 8 (9), 532 (2016).
- Park, H. M., et al. Mass spectrometry-based metabolomic and lipidomic analyses of the effects of dietary Platycodon grandiflorum on liver and serum of obese mice under a high-fat diet. Nutrients. 9 (1), 71 (2017).
- Qi, C., et al. Platycodon grandiflorus polysaccharide with anti-apoptosis, anti-oxidant and anti-inflammatory activity against LPS/D-GalN induced acute liver injury in mice. Journal of Polymers and the Environment. 29 (12), 4088-4097 (2021).
- Choi, J. H., et al. Saponins from the roots of Platycodon grandiflorum ameliorate high fat diet-induced non-alcoholic steatohepatitis. Biomedicine & Pharmacotherapy. 86, 205-212 (2017).
- Choi, Y. J., et al. Platycodin D enhances LDLR expression and LDL uptake via down-regulation of IDOL mRNA in hepatic cells. Scientific Reports. 10, 19834 (2020).
- Li, T., et al. Platycodin D triggers autophagy through activation of extracellular signal-regulated kinase in hepatocellular carcinoma HepG2 cells. European Journal of Pharmacology. 749, 81-88 (2015).
- Lu, J. -. J., et al. Proteomic analysis of hepatocellular carcinoma HepG2 cells treated with platycodin D. Chinese Journal of Natural Medicines. 13 (9), 673-679 (2015).
- Neuschwander-Tetri, B. A. Therapeutic landscape for NAFLD in 2020. Gastroenterology. 158 (7), 1984-1998 (2020).
- Friedman, S. L., Neuschwander-Tetri, B. A., Rinella, M., Sanyal, A. J. Mechanisms of NAFLD development and therapeutic strategies. Nature Medicine. 24 (7), 908-922 (2018).
- Bessone, F., Razori, M. V., Roma, M. G. Molecular pathways of nonalcoholic fatty liver disease development and progression. Cellular and Molecular Life Sciences. 76 (1), 99-128 (2019).
- Buzzetti, E., Pinzani, M., Tsochatzis, E. A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism. 65 (8), 1038-1048 (2016).
- Watt, M. J., Miotto, P. M., De Nardo, W., Montgomery, M. K. The liver as an endocrine organ-Linking NAFLD and insulin resistance. Endocrine Reviews. 40 (5), 1367-1393 (2019).
- Khan, R. S., Bril, F., Cusi, K., Newsome, P. N. Modulation of insulin resistance in nonalcoholic fatty liver disease. Hepatology. 70 (2), 711-724 (2019).
- Karkucinska-Wieckowska, A., et al. Mitochondria, oxidative stress and nonalcoholic fatty liver disease: A complex relationship. European Journal of Clinical Investigation. 52 (3), 13622 (2022).
- Tilg, H., Adolph, T. E., Dudek, M., Knolle, P. Non-alcoholic fatty liver disease: The interplay between metabolism, microbes and immunity. Nature Metabolism. 3 (12), 1596-1607 (2021).
- Qian, H., et al. Autophagy in liver diseases: A review. Molecular Aspects of Medicine. 82, 100973 (2021).
- Du, J., Ji, Y., Qiao, L., Liu, Y., Lin, J. Cellular endo-lysosomal dysfunction in the pathogenesis of non-alcoholic fatty liver disease. Liver International. 40 (2), 271-280 (2020).
- Allaire, M., Rautou, P. E., Codogno, P., Lotersztajn, S. Autophagy in liver diseases: Time for translation. Journal of Hepatology. 70 (5), 985-998 (2019).
- Kanuri, G., Bergheim, I. In vitro and in vivo models of non-alcoholic fatty liver disease (NAFLD). International Journal of Molecular Sciences. 14 (6), 11963-11980 (2013).
- Lau, J. K., Zhang, X., Yu, J. Animal models of non-alcoholic fatty liver disease: Current perspectives and recent advances. The Journal of Pathology. 241 (1), 36-44 (2017).
- Reimer, K. C., Wree, A., Roderburg, C., Tacke, F. New drugs for NAFLD: Lessons from basic models to the clinic. Hepatology International. 14 (1), 8-23 (2020).
- Carpino, G., et al. Increased liver localization of lipopolysaccharides in human and experimental NAFLD. Hepatology. 72 (2), 470-485 (2020).
- Vergani, L. Fatty acids and effects on in vitro and in vivo models of liver steatosis. Current Medicinal Chemistry. 26 (19), 3439-3456 (2019).
- Scorletti, E., Carr, R. M. A new perspective on NAFLD: Focusing on lipid droplets. Journal of Hepatology. 76 (4), 934-945 (2022).
- Green, C. J., Pramfalk, C., Morten, K. J., Hodson, L. From whole body to cellular models of hepatic triglyceride metabolism: Man has got to know his limitations. American Journal of Physiology-Endocrinology and Metabolism. 308 (1), 1-20 (2015).
- Gambino, R., et al. Different serum free fatty acid profiles in NAFLD subjects and healthy controls after oral fat load. International Journal of Molecular Sciences. 17 (4), 479 (2016).
- Marra, F., Svegliati-Baroni, G. Lipotoxicity and the gut-liver axis in NASH pathogenesis. Journal of Hepatology. 68 (2), 280-295 (2018).
- Zhang, J., Zhang, H., Deng, X., Zhang, Y., Xu, K. Baicalin protects AML-12 cells from lipotoxicity via the suppression of ER stress and TXNIP/NLRP3 inflammasome activation. Chemico-Biological Interactions. 278, 189-196 (2017).
- Liang, Y., et al. γ-Linolenic acid prevents lipid metabolism disorder in palmitic acid-treated alpha mouse liver-12 cells by balancing autophagy and apoptosis via the LKB1-AMPK-mTOR pathway. Journal of Agricultural and Food Chemistry. 69 (29), 8257-8267 (2021).
- Peng, Z., et al. Nobiletin alleviates palmitic acid-induced NLRP3 inflammasome activation in a sirtuin 1dependent manner in AML12 cells. Molecular Medicine Reports. 18 (6), 5815-5822 (2018).
- Xu, T., et al. Ferulic acid alleviates lipotoxicity-induced hepatocellular death through the SIRT1-regulated autophagy pathway and independently of AMPK and Akt in AML-12 hepatocytes. Nutrition & Metabolism. 18 (1), 13 (2021).
- Aranda, A., et al. Dichloro-dihydro-fluorescein diacetate (DCFH-DA) assay: A quantitative method for oxidative stress assessment of nanoparticle-treated cells. Toxicology in Vitro. 27 (2), 954-963 (2013).
- Eruslanov, E., Kusmartsev, S. Identification of ROS using oxidized DCFDA and flow-cytometry. Methods in Molecular Biology. 594, 57-72 (2010).
- Bankhead, P. . Analyzing Fluorescence Microscopy Images with ImageJ. , (2014).
- Wiesmann, V., et al. Review of free software tools for image analysis of fluorescence cell micrographs. Journal of Microscopy. 257 (1), 39-53 (2015).
- Lugli, E., Troiano, L., Cossarizza, A. Polychromatic analysis of mitochondrial membrane potential using JC-1. Current Protocols in Cytometry. , (2007).
- Sivandzade, F., Bhalerao, A., Cucullo, L. Analysis of the mitochondrial membrane potential using the cationic JC-1 dye as a sensitive fluorescent probe. Bio-protocol. 9 (1), 3128 (2019).
- Chazotte, B. Labeling mitochondria with JC-1. Cold Spring Harbor Protocols. 2011 (9), (2011).
- Walker, J. M. The bicinchoninic acid (BCA) assay for protein quantitation. The Protein Protocols Handbook. , 11-15 (2009).
- Goldman, A., Ursitti, J. A., Mozdzanowski, J., Speicher, D. W. Electroblotting from polyacrylamide gels. Current Protocols in Protein Science. 82, 1-16 (2015).
- Mozdzanowski, J., Speicher, D. W. Proteins from polyacrylamide gels onto PVDF membranes. Current Research in Protein Chemistry. , 87 (2012).
- Taylor, S. C., Posch, A. The design of a quantitative western blot experiment. Biomed Research International. 2014, 361590 (2014).
- Motulsky, H. J. Graphpad Statistics Guide. Options for multiple t tests. Graphpad. , (2020).
- Poltorak, A. Cell death: All roads lead to mitochondria. Current Biology. 32 (16), 891-894 (2022).
- Dadsena, S., Jenner, A., García-Sáez, A. J. Mitochondrial outer membrane permeabilization at the single molecule level. Cellular and Molecular Life Sciences. 78 (8), 3777-3790 (2021).
- Green, D. R., Kroemer, G. The pathophysiology of mitochondrial cell death. Science. 305 (5684), 626-629 (2004).
- Lange, N. F., Radu, P., Dufour, J. F. Prevention of NAFLD-associated HCC: Role of lifestyle and chemoprevention. Journal of Hepatology. 75 (5), 1217-1227 (2021).
- Liu, X., Zhang, Y., Ma, C., Lin, J., Du, J. Alternate-day fasting alleviates high fat diet induced non-alcoholic fatty liver disease through controlling PPARalpha/Fgf21 signaling. Molecular Biology Reports. 49 (4), 3113-3122 (2022).
- Romero-Gomez, M., Zelber-Sagi, S., Trenell, M. Treatment of NAFLD with diet, physical activity and exercise. Journal of Hepatology. 67 (4), 829-846 (2017).
- Mizushima, N., Levine, B. Autophagy in human diseases. New England Journal of Medicine. 383 (16), 1564-1576 (2020).
- Cui, B., Yu, J. M. Autophagy: A new pathway for traditional Chinese medicine. Journal of Asian Natural Products Research. 20 (1), 14-26 (2018).
- Law, B. Y., et al. New potential pharmacological functions of Chinese herbal medicines via regulation of autophagy. Molecules. 21 (3), 359 (2016).
- Zhou, H., et al. Research progress in use of traditional Chinese medicine monomer for treatment of non-alcoholic fatty liver disease. European Journal of Pharmacology. 898, 173976 (2021).
- Zhang, L., Yao, Z., Ji, G. Herbal extracts and natural products in alleviating non-alcoholic fatty liver disease via activating autophagy. Frontiers in Pharmacology. 9, 1459 (2018).
- Zhang, X., et al. C-X-C motif chemokine 10 impairs autophagy and autolysosome formation in non-alcoholic steatohepatitis. Theranostics. 7 (11), 2822-2836 (2017).
- Li, C. X., et al. Allyl isothiocyanate ameliorates lipid accumulation and inflammation in nonalcoholic fatty liver disease via the Sirt1/AMPK and NF-kappaB signaling pathways. World Journal of Gastroenterology. 25 (34), 5120-5133 (2019).
- Li, S., et al. Sirtuin 3 acts as a negative regulator of autophagy dictating hepatocyte susceptibility to lipotoxicity. Hepatology. 66 (3), 936-952 (2017).
- Farrell, G. C., Teoh, N. C., McCuskey, R. S. Hepatic microcirculation in fatty liver disease. The Anatomical Record. 291 (6), 684-692 (2008).
- Milner, E., et al. Emerging three-dimensional hepatic models in relation to traditional two-dimensional in vitro assays for evaluating drug metabolism and hepatoxicity. Medicine in Drug Discovery. 8, 100060 (2020).
- Zhang, X., Jiang, T., Chen, D., Wang, Q., Zhang, L. W. Three-dimensional liver models: State of the art and their application for hepatotoxicity evaluation. Critical Reviews in Toxicology. 50 (4), 279-309 (2020).
- Bilson, J., Sethi, J. K., Byrne, C. D. Non-alcoholic fatty liver disease: A multi-system disease influenced by ageing and sex, and affected by adipose tissue and intestinal function. Proceedings of the Nutrition Society. 81 (2), 146-161 (2022).
