A neurotransmissão de dopamina (DA) desempenha um papel essencial em várias funções cognitivas e comportamentais, e sua disfunção está implicada em várias doenças e distúrbios comuns. Como tal, é fundamental desenvolver métodos precisos de estudo quantitativo da neurotransmissão DA in vivo para avaliar como a neurotransmissão DA é alterada nos contextos de modelos de doenças e farmacologia de drogas. A voltametria cíclica de varredura rápida (FSCV) permite monitorizar a neurotransmissão DA DA in vivo com resolução espacial e temporal fina. Embora seja possível monitorizar a neurotransmissão de DA fisiológica em animais acordados, que se comportam livremente, a estimulação elétrica de caminhos dopaminérgicos ascendentes em animais anestesiados pode produzir respostas de DA robustas que são favoráveis à análise cinética aprimorada da neurotransmissão DA.
As respostas de DA estimuladas eletronicamente refletem uma interação dinâmica da liberação e recaptação de DA, e interpretaçõesDessas respostas usaram predominantemente um modelo simples de neurotransmissão de DA estimulada chamada modelo Michaelis-Menten (MM) 12 . O modelo de MM consiste em 3 variáveis para descrever as respostas de DA em termos de uma taxa de liberação constante de DA e uma eficiência de recaptação constante ( ou seja, a relação entre a taxa de recaptação de DA e as concentrações extracelulares de DA), conforme descrito pela Equação 1 : 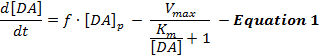
(Liberação de DA) (recaptação de DA)
Na Equação 1, f é a freqüência de estimulação; [DA] p é o aumento estimado da concentração DA por pulso de estimulação; V max representa a taxa de recaptação máxima estimada; E K m é a constante de MM estimada, que é teoricamente equivalente à concentração de DA extracelular que satura 50% de DAT, levando a uma taxa de recaptação de meio máximo. Essa diferençaA equação completa pode ser integrada para simular respostas experimentais de DA estimando os parâmetros [DA] p , V max e K m .
Embora o modelo de MM tenha facilitado avanços significativos na compreensão da cinética de neurotransmissão de DA em vários contextos experimentais, o modelo de MM faz suposições fundamentais simplistas que limitam sua aplicabilidade ao modelar as respostas da DA suscitadas por estimulações suprafisiológicas 2 , 13 . Por exemplo, o modelo de MM só pode aproximar as formas de resposta de DA se aumentar de maneira convexa, mas não pode explicar as respostas crescentes graduais (côncavas) encontradas nas regiões estriadas dorsais 12 . Assim, os pressupostos do modelo MM não capturam com precisão os processos de liberação dinâmica e recaptação da neurotransmissão de DA estimulada.
Para modelar as respostas de DA estimuladas de acordo com um quantum realistaO quadro quantitativo neurobiológico (QN) foi desenvolvido com base em princípios de cinética de neurotransmissão estimulada, derivados de pesquisas complementares e experimentação 2 . Várias linhas de pesquisa de neurotransmissão demonstram que (1) a liberação de neurotransmissor estimulada é um processo dinâmico que diminui na taxa ao longo da estimulação 14 , (2) a liberação continua na fase pós-estimulação com cinética de decomposição bifásica 15 e (3) DA A eficiência da recaptação é progressivamente inibida durante a duração da própria estimulação 2 , 16 . Esses três conceitos servem de base à estrutura do QN e as três equações consistindo em 12 parâmetros que descrevem a dinâmica da liberação e recaptação do DA ( Tabela 1 ). A estrutura de QN pode simular de forma precisa tipos de resposta de DA experimentais heterogêneos, bem como a pEfeitos redicados de manipulações experimentais de parâmetros de estimulação e administração de fármaco 2 , 6 . Embora seja necessária uma pesquisa adicional para refinar a abordagem de modelagem de dados, as experiências futuras podem se beneficiar grandemente dessa abordagem de modelagem neuroquimicamente fundamentada, o que aumenta significativamente as inferências extraídas do paradigma de neurotransmissão DA estimulado.

Tabela 1: Modelagem de Equações e Parâmetros . Clique aqui para ver uma versão maior desta figura.
Este tutorial descreve como modelar dados de resposta de DA estimulados para estimar a liberação de DA e a cinética de recaptação usando o QNsim 1.0. A coleção de dados experimental real e prO processamento não é descrito aqui e requer apenas dados de concentração de DA temporais. O suporte teórico e os fundamentos da estrutura QN foram amplamente descritos anteriormente 2 , mas uma perspectiva prática sobre a aplicação da estrutura QN para o modelo de dados de resposta DA é descrita abaixo.
A estrutura de QN modela a interação dinâmica entre: 1) liberação de DA dinâmica, 2) recaptação de DA e 3) os efeitos de estimulações suprafisiológicas nesses processos para extrair informações cinéticas significativas de dados de resposta de DA. A estrutura QN é mais adequada para modelagem de dados FSCV adquiridos com estimulações altamente suprafisiológicas de longa duração ( por exemplo, 60 Hz, 10 s estimulações), que produzem respostas robustas DA que são passíveis de análise cinética. Seguindo a modelagem precisa dos processos subjacentes de liberação e recaptação, os parâmetros do modelo podem ser usados para simular uma resposta DA que deve se aproximar da forma do exResposta de DA experimental.
As equações da estrutura de QN descrevem as taxas de liberação de DA e recaptação ao longo das respostas de DA estimuladas. A estrutura de QN descreve a taxa de liberação de DA estimulada como uma função do tempo desde o início da estimulação (t stim ), quando a taxa de liberação de DA diminui exponencialmente ao longo da estimulação. Isso é consistente com o esgotamento de um pool prontamente liberável, com uma taxa de liberação de DA de estado estável adicionada (DARss) para explicar o reabastecimento de vesículas, semelhante a outros relatórios ( Equação 2 ) 14 , 17 .
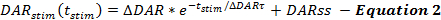
Manipulações que aumentam a taxa de liberação de DA, como o aumento de Δ DAR, Δ DAR τ ou DARss, levam a amplitudes de resposta aumentadas em partos de DA versus tempo. Cada parameE contribui diferencialmente com as formas de resposta DA. Aumentar DARss e Δ DAR τ tornam a fase ascendente das respostas mais linear (menos convexa). Diminuir Δ DAR τ promove a convexidade, que é controlada pela magnitude de Δ DAR. Com base na experiência de modelagem, DARss geralmente é inferior a 1/5 de Δ DAR; Assim, Δ DAR é o parâmetro de lançamento que determina principalmente a amplitude de resposta global de uma resposta DA.
A taxa de liberação de DA pós-estimulação é modelada pela Equação 3 como uma continuação da taxa estimada de liberação de DA desde o final da estimulação (DAR ES ) em função do tempo após a estimulação (t post ). A taxa de liberação de DA pós-estimulação segue um padrão de decaimento bifásico, como descrito anteriormente 15 , com uma rápida fase de decaimento exponencial e uma fase de decaimento linear prolongada para modelar dois caProcessos de liberação de neurotransmissores dependentes de cálcio.
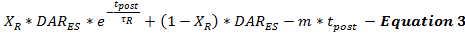
(Decadência exponencial rápida) (Decadência linear prolongada)
Atualmente, não é possível determinar a quantidade de liberação de DA pós-estimulação. Essa limitação pode ser abordada minimizando sistematicamente as estimativas da liberação de DA pós-estimulação e validando os parâmetros do modelo em um conjunto de respostas de DA experimentais coletadas do mesmo local de gravação usando diferentes durações de estimulação. Essa minimização permite aos usuários fazer estimativas conservadoras de liberação e recaptação. Como as estimulações elétricas levam à acumulação de cálcio que promove a liberação pós-estimulação de neurotransmissores, a duração da estimulação influencia os neurotrans pós-estimulaçãoMitter Release parameters 18 , 19 . Com base na experiência de modelagem, verificou-se que, à medida que a duração da estimulação aumenta, τ R aumenta e X R diminui, consistente com os efeitos antecipados de maior acumulação de cálcio 20 .
A Equação 4 descreve a taxa de recaptação de DA como uma extensão da estrutura de MM e incorpora um termo de K m dinâmico, que aumenta durante a estimulação para modelar uma eficiência de recaptação progressivamente decrescente causada pelas estimulações suprafisiológicas 2 , 16 . O K m após a estimulação é mantido constante no valor de K m no final da estimulação (K mES ).
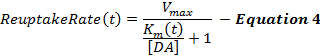
Onde,
<img alT = "Equação 6" src = "/ files / ftp_upload / 55595 / 55595eq6.jpg" />
(Durante a estimulação) (após a estimulação)
As respostas de DA estimuladas, especialmente das regiões estriatares ventrais, são muitas vezes insensíveis às mudanças no valor inicial de K m (K mi ), o que torna a definição de um problema de K mi problemático. Assim, como a estrutura MM original, K mi é aproximado em 0,1-0,4 μ M para respostas DA coletadas de animais não tratados não tratados 12 . O termo Δ K m determina a extensão da mudança de eficiência da recaptação durante a estimulação, que da nossa experiência é de cerca de 2081; M ao longo de uma estimulação de 60 Hz, 10 s. Os valores de K e K minf determinam o modo como K m muda ao longo do tempo e o aumento de cada um destes termos promove a concavidade da fase ascendente. V max é a taxa de recaptação máxima que se relaciona parcialmente com a densidade do transportador DA local, que exibe um gradiente ventromedial para dorsolateral 21 . Por conseguinte, os valores de V max no estriado dorsal (D-Str) são geralmente superiores a 30 μ M / s, mas geralmente inferiores a 30 μ M / s nas regiões ventrais, como o núcleo accumbens (NAc) 6 .
As diretrizes gerais acima podem auxiliar na modelagem de dados experimentais de resposta DA, mas gerar uma simulação que se aproxima da resposta DA experimental precisa ajustar os parâmetros do modelo iterativamente. A precisão dos parâmetros do modelo pode ser melhorada através da obtenção de respostas DA para estimulações suprafisiológicas que proporcionamUm substrato robusto para simulação, bem como através da obtenção e modelagem de múltiplas respostas de DA para estimulações de diferentes durações no mesmo local de gravação ( por exemplo, estimulações de 60 Hz, 5 e 10) para validar a precisão dos parâmetros ( Veja os dados da amostra). Para demonstrar, um conjunto de dados é incluído no pacote de software que contém respostas DA estimuladas regiospecificas coletadas no núcleo accumbens e em estriado dorsal, antes e depois de um desafio farmacológico que já foi modelado usando a estrutura QN. Por extensão, os usuários acharão que esta metodologia pode ser aplicada de forma semelhante para caracterizar a cinética da neurotransmissão DA em vários contextos de doenças e manipulações farmacológicas.
