Дорамин (DA) нейропередача играет существенную роль в различных когнитивных и поведенческих функциях, а ее дисфункция связана с несколькими распространенными заболеваниями и расстройствами. Таким образом, крайне важно разработать точные методы количественного изучения нейротрансмиссии DA in vivo для оценки изменения нейротрансмиссии DA в контексте моделей болезни и фармакологии лекарственных препаратов. Быстрая сканирование циклической вольтамперометрии (FSCV) позволяет осуществлять мониторинг in vivo DA нейротрансмиссии с точным пространственным и временным разрешением. В то время как можно наблюдать физиологическую DA нейротрансмиссию у бодрствующих, свободно ведящих животных, электрическая стимуляция восходящих допаминергических путей у анестезированных животных может приводить к устойчивым ответам ДА, которые поддаются усиленному кинетическому анализу нейротрансмиссии DA.
Электрически стимулированные ответы DA отражают динамическое взаимодействие высвобождения и повторного поглощения DA и интерпретацииИз этих ответов преимущественно использовалась простая модель стимулированной нейротрансмиссии DA, называемая моделью Михаэлиса-Ментена (MM) 12 . Модель ММ состоит из 3 переменных для описания ответов DA в терминах постоянной скорости высвобождения DA и постоянной эффективности обратного захвата ( т. Е. Соотношения между частотой обратного захвата DA и внеклеточными DA-концентрациями), как описано в уравнении 1 : 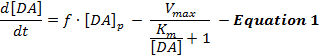
(Выпуск DA) (повторный захват DA)
В уравнении 1 f – частота стимуляции; [DA] p – расчетное увеличение концентрации ДА на импульс стимуляции; V max представляет оценочную максимальную скорость обратного захвата; И K m – оценочная константа ММ, которая теоретически эквивалентна внеклеточной концентрации DA, которая насыщает 50% DAT, что приводит к полумаксимальному коэффициенту обратного захвата. Это отличаетсяAl можно интегрировать для моделирования экспериментальных DA-ответов путем оценки параметров [DA] p , V max и K m .
Хотя модель ММ способствовала значительным достижениям в понимании кинетики нейротрансмиссии DA в различных экспериментальных условиях, модель ММ делает упрощенные фундаментальные предположения, которые ограничивают ее применимость при моделировании ответов ДА, вызванных супрафизиологическими стимуляциями 2 , 13 . Например, модель ММ может только приближать формы ответа ДА, если они растут выпуклым образом, но она не может учитывать постепенные (вогнутые) нарастающие ответы, обнаруженные в дорзальных полосатых областях 12 . Таким образом, предположения модели ММ не точно фиксируют процессы динамического высвобождения и обратного захвата стимулированной нейротрансмиссии DA.
Моделировать стимулированные ответы ДА в соответствии с реалистичным квантом(QN) была разработана на основе принципов стимулированной кинетики нейротрансмиссии, полученных из дополнительных исследований и экспериментов 2 . Различные линии исследований нейротрансмиссии показывают, что (1) стимулированное высвобождение нейротрансмиттера является динамическим процессом, который снижает скорость в течение стимуляции 14 , (2) высвобождение продолжается в фазе после стимуляции с кинетикой бифазного распада 15 и (3) DA Эффективность обратного захвата постепенно ингибируется во время самой стимуляции 2 , 16 . Эти три концепции служат основой структуры QN и трех уравнений, состоящих из 12 параметров, описывающих динамику высвобождения DA и обратного захвата ( таблица 1 ). Структура QN может моделировать гетерогенные экспериментальные типы ответов DA, а также pВыведенные эффекты экспериментальных манипуляций с параметрами стимуляции и введением лекарственного средства 2 , 6 . Хотя дальнейшие исследования необходимы для уточнения подхода к моделированию данных, будущие эксперименты могут в значительной степени выиграть от этого нейробиологически обоснованного подхода к моделированию, что значительно добавляет к выводам, сделанным из стимулированной парадигмы нейротрансмиссии DA.

Таблица 1: Моделирование уравнений и параметров . Нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
В этом учебном пособии описывается, как моделировать стимулированные данные ответа DA для оценки кинетики высвобождения DA и кинетики обратного захвата с использованием QNsim 1.0. Фактический сбор экспериментальных данных и пр.Обработка здесь не описывается и требует только временных данных о концентрации DA. Теоретическая поддержка и основы структуры QN были подробно описаны ранее 2 , но ниже описана практическая перспектива применения структуры QN для моделирования данных ответа DA.
Структура QN моделирует динамическое взаимодействие между: 1) динамическим выпуском DA, 2) обратным захватом DA и 3) эффектами супрафизиологических стимулов на эти процессы для извлечения значимой кинетической информации из данных ответа DA. Структура QN лучше всего подходит для моделирования данных FSCV, полученных с использованием высоко супрафизиологических стимуляций длительной продолжительности ( например, стимуляции 60 Гц, 10 с), которые создают надежные ответы ДА, которые поддаются анализу на кинетику. После точного моделирования основных процессов выпуска и обратного захвата параметры модели могут использоваться для имитации ответа DA, который должен аппроксимировать форму exЭкспериментальный ответ DA.
Уравнения структуры QN описывают скорости высвобождения DA и повторного поглощения в течение стимулированных ответов DA. Структура QN описывает стимулированную скорость высвобождения DA как функцию времени от начала стимуляции (t-стимул), когда скорость высвобождения DA экспоненциально уменьшается в течение стимуляции. Это согласуется с истощением легко освобождаемого пула с добавленной стабильной скоростью высвобождения DA (DARss) для учета пополнения везикул, как и другие отчеты (уравнение 2 ) 14 , 17 .
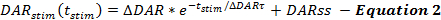
Манипуляции, которые увеличивают скорость высвобождения DA, такие как увеличение ΔDAR, Δ DAR τ или DARss, приводят к увеличению амплитуд ответов на DA по сравнению с временными графиками. Каждый параметрTer вносит дифференциальный характер в формы ответа DA. Увеличение DARss и Δ DAR τ приводит к тому, что нарастающая фаза ответов более линейна (менее выпуклая). Уменьшение Δ DAR τ способствует выпуклости, которая контролируется величиной Δ DAR. Основываясь на опыте моделирования, DARss обычно составляет менее 1/5 Δ ΔDAR; Таким образом, Δ DAR является параметром высвобождения, который в основном определяет общую амплитуду отклика ответа DA.
Скорость высвобождения DA после стимуляции моделируется уравнением 3 в качестве продолжения стимулированной скорости высвобождения DA от конца стимуляции (DAR ES ) в зависимости от времени после стимуляции (t post ). Скорость высвобождения DA после стимуляции следует за двухфазной картиной распада, как описано выше 15 , с быстрой фазой экспоненциального затухания и продолжительной фазой линейного распада для моделирования двух caLcium-зависимых процессов высвобождения нейротрансмиттеров.
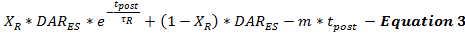
(Быстрое экспоненциальное распад) (Длительный линейный распад)
В настоящее время невозможно определить, сколько возникает пост-стимуляция DA. Это ограничение может быть устранено путем систематического минимизации оценок высвобождения DA после стимуляции и проверки параметров модели по набору экспериментальных ответов ДА, собранных с одного и того же сайта записи, с использованием различных продолжительности стимуляции. Эта минимизация позволяет пользователям делать консервативные оценки выпуска и обратного захвата. Поскольку электрические стимуляции приводят к накоплению кальция, который стимулирует высвобождение нейротрансмиттера после стимуляции, продолжительность стимуляции влияет на постстимулятивные нейротрантыПараметры выхода миттера 18 , 19 . Основываясь на опыте моделирования, было обнаружено, что по мере увеличения продолжительности стимуляции τ R увеличивается, а X R уменьшается, что согласуется с ожидаемым эффектом более высокого накопления кальция 20 .
Уравнение 4 описывает коэффициент обратного захвата DA как расширение каркаса ММ и включает динамический член K m , который увеличивается во время стимуляции, чтобы моделировать постепенно уменьшающуюся эффективность обратного захвата, вызванную супрафизиологическими стимуляциями 2 , 16 . K m после стимуляции поддерживается постоянным при значении K m в конце стимуляции (K mES ).
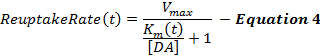
где,
<img alT = "Уравнение 6" src = "/ files / ftp_upload / 55595 / 55595eq6.jpg" />
(Во время стимуляции) (после стимуляции)
Стимулированные ответы ДА, особенно из брюшных полосатых областей, часто нечувствительны к изменениям начального значения K m (K mi ), что делает определение K mi значением проблематичным. Таким образом, как и исходная структура ММ, K mi приближается к 0,1-0,4 мкМ для ответов DA, полученных от контрольных необработанных животных 12 . Термин Δ K m определяет степень изменения эффективности повторного поглощения во время стимуляции, которая по нашему опыту составляет около 2081; M в течение 60-Гц, 10-секундной стимуляции. Значения k и K minf определяют, как K m изменяется с течением времени, и увеличение любого из этих условий способствует вогнутости фазы роста. V max представляет собой максимальную скорость обратного захвата, которая частично относится к локальной плотности транспортера DA, которая проявляет вентромедиальный дорсолатеральный градиент 21 . Соответственно, значения V max в дорсальной полосатой (D-Str) обычно больше 30 мкМ / с, но обычно менее 30 мкМ / с в брюшных областях, таких как ядро accumbens (NAc) 6 .
Общие руководящие принципы, приведенные выше, могут помочь в моделировании экспериментальных данных отклика DA, но генерация моделирования, которая приближается к экспериментальному ответу DA, требует итеративной корректировки параметров модели. Точность параметров модели может быть улучшена путем получения ответов ДА на супрафизиологические стимуляции, которые обеспечиваютEa надежную подложку для моделирования, а также путем получения и моделирования множественных ответов DA на стимуляции различной продолжительности на одном и том же участке записи ( например, стимуляции 60 Гц, 5 и 10 сек) для проверки точности параметров ( См. Примеры данных). Чтобы продемонстрировать, набор данных включен в пакет программного обеспечения, содержащий региоспецифические стимулированные ответы DA, собранные в ядре accumbens и дорсальном полосатом, до и после фармакологической проблемы, которая уже смоделирована с использованием структуры QN. В дополнение, пользователи найдут, что эта методология может быть аналогично применена для характеристики кинетики нейротрансмиссии DA в различных контекстах заболевания и фармакологических манипуляциях.
