Dopamin (DA) neurotransmission spelar en viktig roll i olika kognitiva och beteendemässiga funktioner, och dess dysfunktion är inblandad i flera vanliga sjukdomar och störningar. Som sådan är det kritiskt att utveckla exakta metoder för att kvantitativt studera DA-neurotransmission in vivo för att utvärdera hur DA-neurotransmission förändras i samband med sjukdomsmodeller och läkemedelsfarmakologi. Snabb-skanning cyklisk voltammetri (FSCV) möjliggör övervakning in vivo DA-neurotransmission med fin spatial och tidsmässig upplösning. Medan det är möjligt att övervaka fysiologisk DA-neurotransmission i vakna, fritt uppfostrade djur kan den elektriska stimuleringen av stigande dopaminerga vägar i bedövade djur producera robusta DA-svar som är mottagliga för den förbättrade kinetiska analysen av DA-neurotransmission.
Elektriskt stimulerade DA-svar speglar ett dynamiskt samspel av DA-frisläppande och återupptagning och tolkningarAv dessa svar har främst använt en enkel modell av stimulerad DA-neurotransmission, kallad Michaelis-Menten (MM) -modellen 12 . MM-modellen består av 3 variabler för att beskriva DA-responser i form av en konstant DA-frisättningshastighet och en konstant återupptagningseffektivitet ( dvs. förhållandet mellan DA-upptagningshastigheten och extracellulära DA-koncentrationer), såsom beskrivs av ekvation 1 : 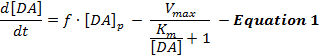
(DA release) (DA återupptagning)
I ekvation 1 är f frekvensen för stimulering; [DA] p är den uppskattade DA-koncentrationens ökning per stimuleringsimpuls; V max representerar den uppskattade maximal återupptagningshastigheten; Och Km är den uppskattade MM-konstanten, som är teoretiskt ekvivalent med den extracellulära DA-koncentrationen som mättar 50% av DAT, vilket leder till en halv maximal återupptagningshastighet. Denna differentiAl ekvation kan integreras för att simulera experimentella DA-svar genom att uppskatta [DA] p , V max och K m parametrar.
Även om MM-modellen har underlättat betydande framsteg i förståelsen av DA-neurotransmissionskinetik i olika experimentella sammanhang, gör MM-modellen enkla grundläggande antaganden som begränsar dess tillämplighet vid modellering DA-svar som framkallas av suprafysiologiska stimulanser 2 , 13 . Exempelvis kan MM-modellen bara approximera DA-responsformer om de stiger på ett konvex sätt, men det kan inte ta hänsyn till de gradvisa (konkava) stigande svaren som finns i dorsala striatala regioner 12 . Således tar MM-modellen antaganden inte exakt upp de dynamiska frisättnings- och återupptagsprocesserna för stimulerad DA-neurotransmission.
Att modellera stimulerade DA-svar enligt en realistisk kvantItative ramverk, den kvantitativa neurobiologiska (QN) ramen utvecklades baserat på principer för stimulerad neurotransmissionskinetik härrörande från komplementär forskning och experiment 2 . Olika linjer av neurotransmissionsforskning visar att (1) stimulerad frigörande av neurotransmitter är en dynamisk process som minskar i takt under stimulans 14 , (2) frisättning fortsätter i post-stimuleringsfasen med bifasisk sönderfallskinetik 15 och (3) DA Reuptake-effektiviteten hämmas progressivt under själva stimulansperioden 2 , 16 . Dessa tre begrepp fungerar som grunden för QN-ramen, och de tre ekvationerna består av 12 parametrar som beskriver dynamiken för DA-frisättning och återupptagning ( Tabell 1 ). QN-ramen kan noggrant simulera heterogena experimentella DA-svarstyper, såväl som pReducerade effekter av experimentella manipuleringar av stimuleringsparametrar och läkemedelsadministration 2 , 6 . Även om ytterligare forskning är nödvändig för att fördjupa datamodelleringsmetoden, kan framtida experiment i stor utsträckning dra nytta av detta neurobiologiskt grundade modellerings-tillvägagångssätt, vilket väsentligt bidrar till de härledda resultaten från det stimulerade DA-neurotransmissionsparadigmet.

Tabell 1: Modelleringsekvationer och parametrar . Vänligen klicka här för att se en större version av denna figur.
Denna handledning beskriver hur man modellerar stimulerad DA-responsdata för att uppskatta DA-frisättning och återupptagskinetik med QNsim 1.0. Den faktiska experimentella datainsamlingen och prUppgift är inte beskrivet här och kräver endast tidsmässig DA-koncentrationsdata. Det teoretiska stödet och grunden för QN-ramen har beskrivits utförligt tidigare 2 , men ett praktiskt perspektiv på att tillämpa QN-ramverket för modell DA-svardata beskrivs nedan.
QN-ramen modellerar det dynamiska samspelet mellan: 1) dynamisk DA-frisättning, 2) DA-återupptagning, och 3) effekterna av suprafysiologiska stimulanser på dessa processer för att extrahera meningsfull kinetisk information från DA-svardata. QN-ramverket passar bäst för modellering av FSCV-data som erhållits med högt suprafysiologiska stimulanser med lång varaktighet ( t.ex. 60 Hz, 10 s stimulanser), vilket ger robusta DA-svar som är mottagliga för kinetisk analys. Efter noggrann modellering av de underliggande frisättnings- och återupptagsprocesserna kan modellparametrarna användas för att simulera ett DA-svar som skulle approximera formen på exPerimental DA svar.
Ekvationerna i QN-ramen beskriver hastigheterna för DA-frisättning och återupptagning under de stimulerade DA-svarenas lopp. QN-ramen beskriver den stimulerade DA-frisättningsgraden som en funktion av tiden från stimulansstart (t-st), när DA-frisättningsgraden exponentiellt minskar under stimulansförloppet. Detta överensstämmer med uttömningen av en lätt frigörbar pool, med en tillsatt stabiliserad DA-frisättningshastighet (DARss) för redovisning av vesikelpåfyllning, som liknar andra rapporter ( ekvation 2 ) 14 , 17 .
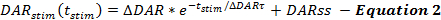
Manipuleringar som ökar DA-frisättningsfrekvensen, såsom ökande ΔDAR, ΔDAR τ eller DARss, leder till ökade responsförstärkningar på DA-versus tidstimer. Varje parameTer bidrar olika till DA-responsformer. Ökande DARss och ΔDAR τ gör båda stigande faser av svar mer linjära (mindre konvexa). Att minska Δ DAR τ främjar konvexitet, som styrs av storleken av Δ DAR. Baserat på modelleringserfarenhet är DARss generellt mindre än 1/5 av Δ DAR; Sålunda är Δ DAR frisättningsparametern som primärt bestämmer den totala responsamplituden för ett DA-svar.
DA-frisättningshastigheten efter stimulering modelleras av ekvation 3 som en fortsättning på den stimulerade DA-frisättningshastigheten från slutet av stimulering (DAR ES ) som en funktion av tiden efter stimulering (t- post ). Post-stimulering DA-frisättningshastigheten följer ett bifasiskt sönderfallsmönster, som tidigare beskrivits 15 , med en snabb exponentiell sönderfallsfas och en förlängd linjär sönderfallsfas till modell två caLciumberoende neurotransmittor-frisättningsförfaranden.
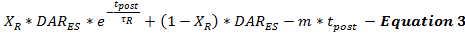
(Snabb exponentiell sönderfall) (långvarigt linjärt förfall)
Det är för närvarande inte möjligt att bestämma hur mycket poststimulering DA-frisättning uppstår. Denna begränsning kan åtgärdas genom att systematiskt minimera uppskattningar av poststimulering DA-frisättning och validering av modellparametrar över en uppsättning experimentella DA-svar som samlas in från samma inspelningsplats med användning av varierande stimuleringstider. Denna minimering tillåter användare att göra konservativa uppskattningar av frisättning och återupptagning. Eftersom elektriska stimuleringar leder till kalciumackumulering som främjar frisättning av neurostransmitter efter stimulering påverkar stimuleringstiden poststimulationsneurotransMitterfrigöringsparametrar 18 , 19 . Baserat på modelleringserfarenhet visade sig att när stimulansvaraktigheten ökar ökar τ R och X R minskar, vilket överensstämmer med de förväntade effekterna av en större kalciumackumulering 20 .
Ekvation 4 beskriver DA-återupptagningsgraden som en förlängning av MM-ramen och införlivar en dynamisk Km-term som ökar under stimulering för att modellera en gradvis minskande återupptagningseffektivitet orsakad av de suprafysiologiska stimulanserna 2 , 16 . Km efter stimulering hålls konstant vid Km-värdet vid slutet av stimuleringen (K mES ).
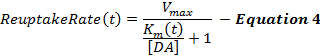
var,
<img alT = "ekvation 6" src = "/ files / ftp_upload / 55595 / 55595eq6.jpg" />
(Under stimulering) (Efter stimulering)
Stimulerade DA-svar, särskilt från ventralstriatala regioner, är ofta okänsliga för förändringar i det ursprungliga Km-värdet (K mi ), vilket gör att ett K mi- värde problematiskt definieras. Således är, som den ursprungliga MM-ramen, K mi approximerad vid 0,1-0,4 μM för DA-svar uppsamlade från kontrollbehandlade djur 12 . Termen Δ K m bestämmer omfattningen av upptagningseffektivitetsförändringen under stimulering, vilket från vår erfarenhet är cirka 2081; M under loppet av en 60 Hz, 10 s stimulering. K- och K- minf- värdena bestämmer hur Km ändras över tiden, och ökning av någon av dessa termer främjar uppgången i stigningsfasen. V max är den maximala återupptagningsgraden som delvis hänför sig till lokal DA-transportörens densitet, vilken uppvisar en ventromedial till dorsolateral gradient 21 . Följaktligen är Vmax- värden i dorsalstriatumet (D-Str) i allmänhet större än 30 ^ M / s men i allmänhet mindre än 30 ^ M / s i de ventrala regionerna, som kärnan accumbens (NAc) 6 .
De allmänna riktlinjerna ovan kan hjälpa till med modellering av experimentella DA-svardata, men generering av en simulering som approximerar det experimentella DA-svaret kräver iterativt att justera modellparametrar. Noggrannheten hos modellparametrarna kan förbättras genom att erhålla DA-svar på suprafysiologiska stimuleringar som tillhandahållerEa robust substrat för simulering, samt genom att erhålla och modellera flera DA-svar på stimuleringar av varierande varaktighet på samma inspelningsplats ( t.ex. stimulering av 60 Hz, 5 s och 10 s) för att validera parameterns noggrannhet ( Se provdata). För att visa att en dataset ingår i mjukvarupaketet som innehåller regiospecifika stimulerade DA-svar som samlats in i kärnans accumbens och dorsalstriatum före och efter en farmakologisk utmaning som redan modellerades med hjälp av QN-ramverket. Utvidgningen kommer användarna att finna att denna metodik kan användas för att karakterisera kinetiken för DA-neurotransmission i olika sjukdomskontext och farmakologiska manipuleringar.
