Planteassocierede mikrober spiller vigtige roller inden for biogeokemisk cykling, bioremediering, afbødning af klimaændringer, plantevækst og sundhed og plantetolerance overfor biotiske og abiotiske påvirkninger. Mikroorganismer interagerer med planter både direkte gennem plantecellevægskontakt og indirekte via kemisk sekretion og signalering 1 , 2 , 3 . Som sessile organismer har planter udviklet direkte og indirekte mekanismer til at modstå infektion med patogener. Direkte forsvar omfatter strukturelle forsvar og udtryk for forsvarsproteiner, mens indirekte forsvar omfatter sekundær plantemetabolitproduktion og tiltrækningen af organismer, der er antagonistiske for invaderende patogener 4 , 5 . Planteafledte rod-exudater, sekreter, slimhinder, mucigel og lysater ændrer rhizosfærens fysisk-kemiske egenskaber for at tiltrække eller afviseMikrober mod deres værter 6 . Den kemiske sammensætning af rodsekretion er artsspecifik og tjener derved som et selektivt filter, der tillader visse mikroorganismer, der er i stand til at genkende sådanne forbindelser for at blomstre i rhizosfæren 6 . Således kan kompatible mikrobielle arter stimuleres til at aktivere og forbedre deres foreninger, enten til fordel for eller skade for planteværten 1 .
Forståelse af plante-mikrobe interaktioner i rhizosfæren er nøglen til at forbedre planteproduktiviteten og økosystemets funktion, da størstedelen af den mikrobielle og kemiske eksponering forekommer ved rodstrukturen og jord-luftgrænsefladen 2 , 6 , 7 , 8 . Undersøgelsen af underjordiske plante-mikrobe-interaktioner og gensidige reaktioner har imidlertid været en udfordring på grund af dens fascinerende Kompleks og dynamisk natur og mangel på egnede eksperimentelle modeller med naturlig rodstruktur og plantemorfologi under tæt kontrollerbare vækstbetingelser. Som en af de mest undersøgte fytopathogener inficerer Agrobacterium en bred vifte af planter med landbrugs- og gartnerisk betydning, herunder kirsebær, æble, pære, drue og rose 9 . Agrobacterium er en vigtig modelorganisme til forståelse af plantepatogeninteraktioner og er et kraftfuldt værktøj i plantetransformation og planteteknik 10 , 11 , 12 , 13 , 14 .
Molekylære plante- Agrobacterium- interaktioner er blevet undersøgt godt i flere årtier, og den nuværende forståelse af Agrobacterium- patogenicitet er omfattende 9 ,F.> 11 , 15 , 16. Agrobacteriumpatogenicitet er i høj grad tilskrevet dets evoluerede evner til at opfatte planteafledte signaler, hvilket resulterer i den fine modulering af dets virulensprogram og celle til celle kommunikation, såkaldt quorum sensing 17 . Agrobacterium virulence-programmet reguleres af flere signaler til rådighed i rhizosfæren og involverer to sæt 2-komponentsystemer, ChvG / I-systemet og VirA / G-systemet. Syrebetingelser i rhizosfæren aktiverer transkriptionen af chvG / I , virA / G , Og flere andre gener involveret i Agrobacterium- patogenicitet, herunder virE0 , virE1 , virH1 , virH2 og gener af type VI-sekretionssystemet (T6SS) 18. Plantafledte phenolforbindelser, herunder acetosyringon (4'-hydroxy-3 ', 5 '-dimethoxyacetophenon), aktiver V'enIrA / G 2-komponentsystem gennem phosphoryleringssignaleringsmekanismer 19 . VirA / G aktiverer derefter hele vir- regulonet, hvilket resulterer i overførsel og integration af et ~ 20 kb bakterielt DNA-fragment kaldet transfer DNA (T-DNA) fra dets tumorinducerende (Ti) plasmid i plantekernen 16 . T-DNA bærer gener ansvarlig for syntesen af plantehormonerne indol-3-eddikesyre (IAA) ( iaaM og iaaH ) og cytokinin ( ipt ) og en gang udtrykt i planteceller produceres store mængder af disse phytohormoner. Dette resulterer i unormal vævsproliferation og udvikling af plantetumor, kendt som kronesygdom, hvilket er et kronisk og genopståeligt problem for planterne 9 , 11 , 20 . IAA virker også kollektivt med salicylsyre og gamma-aminobutyrsyre for at undertrykke Agrobacterium virulens eller for at reducere Agrobacteriu M quorum sensing (QS) 17 , 21 , 22 . For at imødegå denne undertrykkelse bærer T-DNA også gener for opinbiosyntese, som aktiverer Agrobacterium quorum-sensing for at fremme Agrobacterium- patogenicitet og tjener også som en næringsstofkilde for patogenet 22 , 23 .
På trods af en samlet dyb forståelse af Agrobacterium- planteinteraktioner og den resulterende T-DNA-overførsel til planteværten er de komplekse signaleringshændelser i den indledende fase af interaktionen mindre velkendte. Dette skyldes delvist begrænsningerne af konventionelle metoder til undersøgelse af Agrobacterium- plantesignalering. Plantecellesuspensionskulturer og kunstig stedsspecifik såring anvendes almindeligt til at studere molekylære plante-mikrobe interaktioner 24 ,Ef "> 26 , 27. Celleopslæmninger mangler imidlertid typisk plantemorfologi, især plantesuspensionsceller har ikke rotstrukturer og rodeksudater, som er meget vigtige for aktivering af mikrobiell kemotaxis og virulens 28 , 29. Vedligeholdelsen af plantemorfologi Og rodstruktur er blevet behandlet af kunstigt sårende planter, hvilket letter lokalitetsspecifik infektion, hvilket resulterer i påvisning af inducerede planteforsvarrelaterede gener i direkte inficeret plantevæv 30 , 31. Imidlertid er kunstig såring signifikant forskellig fra patogeninfektion i naturen , Især da sår fører til akkumulering af jasmoninsyre (JA), som systematisk interfererer med naturlig plantesignalering og forsvar 26. Desuden anvendes syntetiske kemikalier typisk til kunstigt at fremkalde planteværtsresponserEller patogen virulens. Skønt tilskuddet af sådanne kemiske forbindelser, der reflekterer koncentrationer i planta, er muligt, tager sådanne tilskud ikke hensyn til diffusionen af rod-exudater gradvist ind i den omgivende rhizosfære, hvilket frembringer en kemotaktisk gradient registreret af mikrober 28 , 32 . I betragtning af begrænsningerne af konventionelle fremgangsmåder til undersøgelse af plante-mikrobe-interaktioner kan nøjagtigheden og dybden af de opnåede data være hæmmet og restriktive, og den viden, der genereres ud fra de konventionelle fremgangsmåder, kan ikke oversættes direkte i planta . Mange aspekter af plante- Agrobacterium- signalering er endnu ikke fuldt ud forstået, især i det tidlige stadium af interaktioner, når sygdomssymptomerne endnu ikke er udviklet.
For at ændre begrænsningerne ved konventionelle tilgange præsenterer dette arbejde en billig, tæt kontrollerbar og fleksibel hydroponisk cOdlingssystem, der giver forskere mulighed for at få dybere indsigt i de komplekse signal- og reaktionsveje ved begyndelsen af molekylære plante-mikrobe interaktioner. Hydroponics har været meget anvendt til at studere plantenæringsstoffer, rod-ekssudater, vækstbetingelser og virkningerne af metallisk toksicitet på planterne 33 , 34 . Der er flere fordele ved hydroponiske modeller, herunder de små rumlige krav, tilgængeligheden af forskellige plantevæv, den tætte kontrol af næringsstof / miljøforhold og bekæmpelse af skadegørere / sygdomme. Hydroponiske systemer er også mindre begrænsende for plantevækst i sammenligning med agar / phytoagar plating teknikker, der typisk begrænser vækst efter 2-3 uger. Det er vigtigt, at vedligeholdelsen af helplantekonstruktioner letter den naturlige rodsekretion, der er nødvendig for mikrobiell kemotakse og virulensinduktion 8 , 29 . Systemet beskriverSeng her er enklere og mindre arbejdskrævende end alternativene 33 , 34 . Det bruger færre dele og kræver ikke andre værktøjer end standard saks. Den bruger metalnet (i modsætning til nylon 33 ) som en stærk støtte til plantevækst og en simpel metode til beluftning under sterile betingelser gennem omrystning for at understøtte mikrobiel vækst. Derudover kan systemet bruge metalnet af forskellige størrelser til at understøtte plantevækst, som rummer forskellige plantearter uden at begrænse bredden af deres rødder.
I det hydroponiske kultiveringssystem, der præsenteres her, dyrkes planter i et sterilt hydroponisk system, hvor plantens rødder udskiller organiske forbindelser, der understøtter væksten af podede bakterier. I dette kultiveringssystem suppleres ingen kunstige kemikalier, såsom plantehormoner, forsvarselektronik eller virulensfremkaldende kemikalier, hvilket afspejler den naturlige celleSignalerende homeostase under plante-mikrobe interaktioner. Med dette hydroponiske kultiveringssystem var det muligt at samtidigt bestemme genekspression i Arabidopsis thaliana Col-0 rodvæv efter infektion med Agrobacterium , såvel som aktiveringen af Agrobacterium- gener ved kultivering med Arabidopsis . Det blev yderligere påvist, at dette system er egnet til at studere Agrobacterium- vedhæftning til planterødder såvel som planterodssekretomprofilen ved kultivering (infektion) med Agrobacterium ( Figur 1 ).
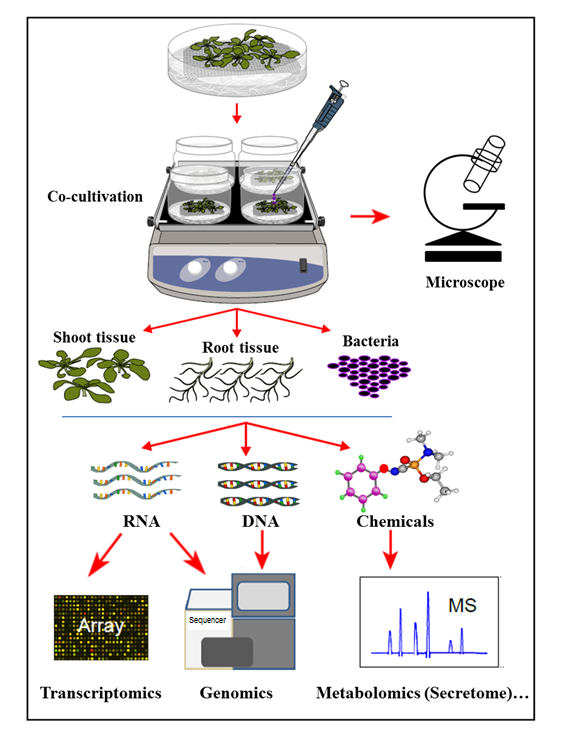
Figur 1: Oversigt over det hydroponiske kultiveringssystem med stikprøveanalyser. Planter dyrkes oven på masken (skud over masken), med rødderne nedsænket i hydroponisk medium, som derefter inokuleres med bakterier fEller kultivering. Plantevæv og bakterier separeres derefter til samtidige ekstraktioner og analyser. Dette tal er blevet ændret fra reference 35 .
