I microbi associati all'impianto svolgono un ruolo importante nel ciclo biogeochimico, nella bioremediazione, nella mitigazione del cambiamento climatico, nella crescita vegetale e nella salute, e nella tolleranza alle tensioni alle tensioni biotiche e abiotiche. I microrganismi interagiscono con le piante sia direttamente attraverso il contatto cellulare a parete cellulare che indirettamente tramite secrezione chimica e segnalazione 1 , 2 , 3 . Come organismi sessili, le piante hanno sviluppato meccanismi diretti e indiretti per resistere all'infezione da agenti patogeni. Le difese dirette comprendono le difese strutturali e l'espressione delle proteine di difesa, mentre le difese indirette includono la produzione secondaria del metabolita vegetale e l'attrazione degli organismi antagonisti agli invasori patogeni 4 , 5 . Gli essudati radici derivanti dalle piante, le secrezioni, le mucillagini, i mucigel e i lisati alterano le proprietà fisico-chimiche della rizosfera per attirare o respingereMicrobi verso i loro ospiti 6 . La composizione chimica della secrezione radicale è specie-specifica, servendo così come un filtro selettivo che consente a certi microrganismi in grado di riconoscere tali composti a fiorire nella rizosfera 6 . Pertanto, le specie microbiche compatibili possono essere stimolate ad attivare e migliorare le loro associazioni, sia a vantaggio che a detrimento dell'ospite vegetale 1 .
La comprensione delle interazioni tra pianta e microbe nella rizosfera è fondamentale per aumentare la produttività delle piante e il funzionamento dell'ecosistema, in quanto la maggior parte delle esposizioni microbiche e chimiche si verifica nella struttura radice e nell'interfaccia del suolo-aria 2 , 6 , 7 , 8 . Tuttavia, l'esame delle interazioni tra piante e micro-piani sotterranee e le risposte reciproche è stata una sfida a causa del suo intrigante Complessa e dinamica e la mancanza di adeguati modelli sperimentali con struttura radicale naturale e morfologia vegetale in condizioni di crescita strettamente controllabili. Come uno dei fitopatogeni più pesantemente studiati, Agrobacterium infetta un'ampia gamma di piante con importanza agricola e orticola, tra cui la ciliegia, la mela, la pera, l'uva e la rosa 9 . Agrobacterium è un organismo di modello importante per la comprensione delle interazioni patogeno-patogeno ed è un potente strumento di trasformazione delle piante e di ingegneria degli impianti 10 , 11 , 12 , 13 , 14 .
Le interazioni molecolari delle piante- Agrobacterium sono state ben studiate per diversi decenni e l'attuale comprensione della patogenicità di Agrobacterium è estesa 9 ,F "> 11 , 15 , 16. La patogenicità di Agrobacterium è in gran parte attribuita alle sue capacità evolute di percepire i segnali derivanti dalle piante, con conseguente fine modulazione del suo programma di virulenza e comunicazione cellulare da cellule, il cosiddetto sensore quorum 17 . Il programma di virulenza di Agrobacterium è regolato da diversi segnali disponibili nella rizosfera e coinvolge due set di sistemi a 2 componenti, il sistema ChvG / I e il sistema VirA / G. Le condizioni acide nella rizosfera attivano la trascrizione di chvG / I , virA / G , E molti altri geni coinvolti nella patogenicità di Agrobacterium , compresi virE0 , virE1 , virH1 , virH2 e geni del sistema di secrezione di tipo VI (T6SS) 18 . '-dimetossiacetofenone), attivare la VSistema 2-componente irA / G attraverso i meccanismi di segnalazione della fosforilazione 19 . VirA / G quindi attiva l'intera regolazione vir , con conseguente trasferimento e integrazione di un frammento di DNA batterico ~ 20 kb chiamato DNA di trasferimento (T-DNA) dal suo tumore-inducente (Ti) plasmide nel nucleo di pianta 16 . T-DNA porta i geni responsabili della sintesi degli ormoni vegetali indole-3-acetico (IAA) ( iaaM e iaaH ) e citochinina ( ipt ) e una volta espressi in cellule vegetali, si producono grandi quantità di questi fitotomi. Ciò si traduce in una proliferazione anomala del tessuto e nello sviluppo del tumore delle piante, noto come malattia della corona, che è un problema cronico e risorgente per le piante 9 , 11 , 20 . L'IAA agisce anche collettivamente con acido salicilico e acido gamma-ammino butirrico per reprimere la virulenza di Agrobacterium o per ridurre l' Agrobacteriu M quorum sensing (QS) 17 , 21 , 22 . Per contrastare questa repressione, T-DNA porta anche dei geni per la biosintesi opina, che attiva Agrobacterium quorum sensing per promuovere la patogenicità di Agrobacterium e serve anche come fonte nutritiva per il patogeno 22 , 23 .
Nonostante una comprensione approfondita delle interazioni di Agrobacterium- Plant e del trasferimento di T-DNA risultante nell'ospite della pianta, gli eventi complessi di segnalazione nella fase iniziale dell'interazione sono meno ben compresi. Ciò è dovuto in parte alle limitazioni degli approcci convenzionali per indagare la segnalazione Agrobacterium- Plant. Le colture di sospensione delle cellule vegetali e ferite artificiali specifiche del sito vengono comunemente utilizzate per studiare le interazioni tra microbi e piante molecolari 24 ,. ef "> 26, 27 Tuttavia, sospensioni cellulari mancare tipica morfologia delle piante, in particolare celle di sospensione pianta non hanno strutture radicolari e essudati radicali, che sono molto importanti per l'attivazione chemiotassi microbica e virulenza 28, 29 Il mantenimento della morfologia delle piante. E la struttura delle radici è stata affrontata da piante ferite artificiali che facilitano l'infezione specifica del sito, con conseguente individuazione di geni correlati alla difesa vegetale indotta nel tessuto vegetale direttamente infetto 30 , 31. Tuttavia, ferita artificiale è significativamente diversa dall'infezione patogena in natura , In particolare in quanto ferita porta ad accumulo di acido jasmone (JA) che interferisce sistematicamente con la segnalazione e la difesa naturale delle piante 26. Inoltre, sono tipicamente impiegate sostanze chimiche sintetiche per indurre artificialmente le risposte dell'organismo vegetaleO la virulenza del patogeno. Anche se l'integrazione di tali composti chimici che riflettono concentrazioni in planta è possibile, tale integrazione non rende conto della diffusione di essudati radici gradualmente nella rizosfera circostante, che genera un gradiente chemotattico rilevato dai microbi 28 , 32 . Data la limitazione degli approcci convenzionali per studiare le interazioni tra pianta e microbe, l'accuratezza e la profondità dei dati ottenuti potrebbero essere ostacolati e restrittivi e le conoscenze generate dagli approcci convenzionali non possono essere tradotte direttamente in planta . Molti aspetti della segnalazione di pianta- Agrobacterium non sono ancora pienamente compresi, in particolare nella fase iniziale delle interazioni, quando i sintomi della malattia non sono ancora stati sviluppati.
Per modificare le limitazioni degli approcci convenzionali, questo lavoro presenta un sistema idroponico poco costoso, strettamente controllabile e flessibileSistema di ocultivazione che consente ai ricercatori di acquisire approfondimenti sui complessi percorsi di segnalazione e risposta nella fase iniziale delle interazioni molecolari delle piante-microbe. L'idroponica è stata ampiamente usata per studiare le sostanze nutritive vegetali, gli essudati radici, le condizioni di crescita e gli effetti della tossicità metallica sulle piante 33 , 34 . Ci sono diversi vantaggi di modelli idroponici, tra cui i piccoli requisiti spaziali, l'accessibilità di vari tessuti vegetali, il controllo stretto delle condizioni nutrizionali / ambientali e il controllo di parassiti / malattie. I sistemi idroponici sono anche meno limitanti per la crescita delle piante rispetto alle tecniche di placcatura agar / phytoagar, che in genere limitano la crescita dopo 2-3 settimane. Importante, il mantenimento delle strutture vegetali intere facilita la secrezione naturale della radice necessaria per la chemiotassi microbica e l'induzione della virulenza 8 , 29 . Il sistema descriIl letto qui è più semplice e meno intensivo rispetto alle alternative 33 , 34 . Utilizza meno parti e non richiede strumenti diversi dalle forbici standard. Utilizza la maglia metallica (in contrapposizione al nylon 33 ) come un forte supporto per la crescita delle piante e un semplice metodo di aerazione in condizioni sterili mediante agitazione per supportare la crescita microbica. Inoltre, il sistema può utilizzare maglie metalliche di varie dimensioni per sostenere la crescita vegetale, che accoglie diverse specie vegetali senza limitare la larghezza delle loro radici.
Nel sistema di coltivazione idroponica qui presentato, le piante vengono coltivate in un sistema idroponico sterile dove le radici vegetali secernono composti organici che sostengono la crescita dei batteri inoculati. In questo sistema di cocultivazione non vengono aggiunti prodotti chimici artificiali, come ormoni vegetali, elicitor di difesa o sostanze chimiche che inducono la virulenza, che riflette la cella naturale-signostasi dell'omeostasi durante le interazioni plant-microbe. Con questo sistema di cocultivazione idroponica è stato possibile determinare simultaneamente l'espressione genica nel tessuto radicale Arabidopsis thaliana Col-0 su infezione da Agrobacterium , nonché l'attivazione di geni Agrobacterium dopo cocultivazione con Arabidopsis . È stato inoltre dimostrato che questo sistema è idoneo a studiare l'attaccamento di Agrobacterium alle radici vegetali, nonché al profilo di segreto radicale di pianta, dopo la cocultivazione (infezione) con Agrobacterium ( Figura 1 ).
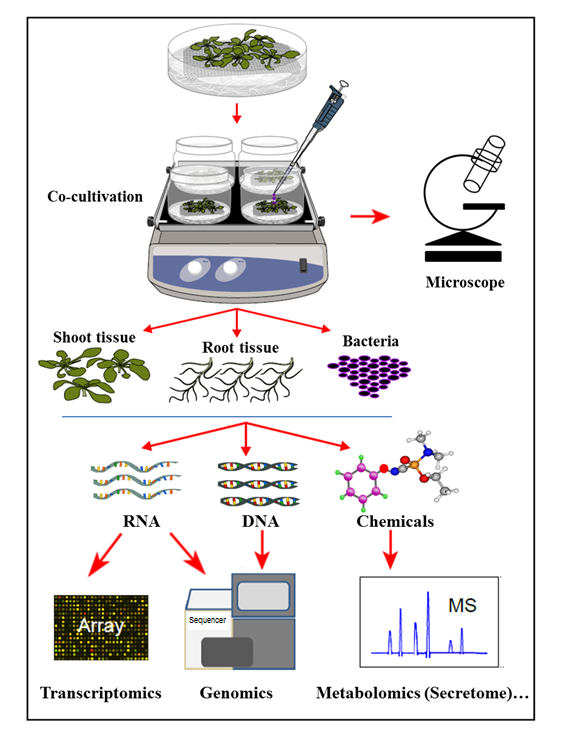
Figura 1: Panoramica del Sistema di Cocultivazione Idroponica, con analisi del campione. Le piante sono coltivate in cima alla maglia (germogli al di sopra della maglia), con le radici immerse in mezzo idroponico che viene quindi inoculato con batteri fO cocultura. I tessuti e i batteri delle piante vengono quindi separati per le estrazioni e le analisi simultanee. Questa cifra è stata modificata dal riferimento 35 .
