Plantgerelateerde microben spelen belangrijke rollen in biogeochemische cycli, bioremediëring, vermindering van klimaatverandering, plantengroei en gezondheid en plantentolerantie tegen biotische en abiotische stress. Micro-organismen interageren met planten, zowel direct door plantencelwandcontact en indirect via chemische afscheiding en signalering 1 , 2 , 3 . Als sessiele organismen hebben planten directe en indirecte mechanismen ontwikkeld om infectie door pathogenen te weerstaan. Directe verdediging omvat structurele verdediging en de expressie van verdedigingseiwitten, terwijl indirecte verdediging de productie van secundaire plantenmetabolieten omvat en de aantrekkingskracht van organismen antagonistisch tegen invallende pathogenen 4 , 5 . Plantaardige wortel exudaten, afscheidingen, mucilages, mucigel en lysaten veranderen de fysisch-chemische eigenschappen van de rhizosfeer om te trekken of af te kerenMicroben naar hun gastheren 6 . De chemische samenstelling van wortelafscheiding is speciesspecifiek, en dient daarbij als selectief filter dat bepaalde micro-organismen in staat stelt om dergelijke verbindingen te herkennen om in de rhizosfeer 6 te bloeien. Zo kunnen compatibele microbiële soorten worden gestimuleerd om hun associaties te activeren en te verbeteren, hetzij tot voordeel of nadelen van de plantengas 1 .
Het begrijpen van plant-microbe-interacties in de rhizosfeer is essentieel voor het verbeteren van de productiviteit van het plant en het functioneren van het ecosysteem, aangezien een meerderheid van de microbiële en chemische blootstelling zich voordoet bij de wortelstructuur en de bodem-luchtinterface 2 , 6 , 7 , 8 . Het onderzoek naar ondergrondse plant-microbe interacties en wederzijdse reacties is echter een uitdaging als gevolg van het intrigerende Complexe en dynamische natuur en het ontbreken van geschikte experimentele modellen met natuurlijke wortelstructuur en plantmorfologie onder strak beheersbare groeivoorwaarden. Als een van de meest bestudeerde fytopathogenen infecteert Agrobacterium een breed scala aan planten met agrarisch en tuinbouw belang, waaronder kersen, appel, peer, druif en roos 9 . Agrobacterium is een belangrijk model organisme voor het interpreteren van plant-pathogeen interacties en is een krachtig instrument in plantentransformatie en planttechniek 10 , 11 , 12 , 13 , 14 .
Moleculaire plant- Agrobacterium interacties zijn al jarenlang bestudeerd en het huidige begrip van Agrobacterium pathogeniciteit is uitgebreid 9 ,F.> 11 , 15 , 16. De pathogeniciteit van Agrobacterium wordt grotendeels toegeschreven aan de evoluerende mogelijkheden van het opnemen van plantaardige afgeleid signalen, wat resulteert in de fijne modulatie van het virulentieprogramma en de cel-naar-celcommunicatie, zogenaamd quorum sensing 17 . Het Agrobacterium virulence programma wordt geregeld door verschillende signalen die beschikbaar zijn in de rhizosfeer en omvatten twee sets van 2-componenten systemen, het ChvG / I-systeem en het VirA / G-systeem. Acide condities in de rhizosfeer activeren de transcriptie van chvG / I , virA / G , En verscheidene andere genen die betrokken zijn bij Agrobacterium pathogeniciteit, waaronder virE0 , virE1 , virH1 , virH2 en genen van het type VI-afzettingssysteem (T6SS) 18. Plantafledde fenolverbindingen, waaronder acetosyringon (4'-hydroxy-3 ', 5 '-dimethoxyacetofenon), activeer de VIrA / G 2-componenten systeem via fosforylatie signaal mechanismen 19 . VirA / G activeert dan het volledige vir- regulon, wat resulteert in de overdracht en integratie van een ~ 20 kb bacterieel DNA fragment genaamd transfer DNA (T-DNA) van zijn tumorinducerende (Ti) plasmide in de plantencel 16 . T-DNA draagt genen die verantwoordelijk zijn voor de synthese van plantaardige hormonen indool-3-azijnzuur (IAA) ( iaaM en iaaH ) en cytokinine ( ipt ), en eenmaal uitgedrukt in plantencellen worden grote hoeveelheden van deze fytohormonen geproduceerd. Dit resulteert in abnormale weefsel proliferatie en ontwikkeling van plantentumor, bekend als kroon galziekte, die een chronisch en resurgent probleem is voor planten 9 , 11 , 20 . IAA handelt ook collectief met salicylzuur en gamma-aminoboterzuur om de Agrobacterium virulentie te onderdrukken of om Agrobacteriu te verminderen M kworum sensing (QS) 17 , 21 , 22 . Om deze onderdrukking tegen te gaan, draagt T-DNA ook genen voor opine biosynthese, die Agrobacterium quorum detecteert om de pathogeniteit van Agrobacterium te bevorderen en dient ook als voedingsbron voor het pathogeen 22 , 23 .
Ondanks een algemeen diepgaand begrip van Agrobacterium- plantinteracties en de resulterende T-DNA-overdracht in de plantaardige gastheer, zijn de complexe signaleringsgebeurtenissen in het beginstadium van interactie minder goed begrepen. Dit komt gedeeltelijk door de beperkingen van conventionele benaderingen om Agrobacterium- plantensignaal te onderzoeken. Plantcelsuspensieculturen en kunstmatige site-specifieke wonden worden vaak gebruikt voor het bestuderen van moleculaire plant-microbe-interacties 24 ,Ef,> 26 , 27. Echter, cel suspensies hebben typische plantmorfologie, in het bijzonder hebben plantaardige suspensiecellen geen wortelstructuren en wortel exudaten, die zeer belangrijk zijn voor het activeren van microbiële chemotaxis en virulentie 28 , 29. Het behoud van plantmorfologie En wortelstructuur is aangepakt door kunstmatig wondende planten, waardoor plaatselijke specifieke infectie wordt vergemakkelijkt, wat resulteert in het detecteren van geïnduceerde plantenverdediging gerelateerde genen in direct geïnfecteerd plantweefsel 30 , 31. Maar kunstmatige wonden is significant verschillend van pathogeeninfectie in de natuur , Met name omdat wonden leidt tot accumulatie van jasmonzuur (JA), die systematisch inbreuk maakt op natuurlijke plantensignalen en verdediging 26. Bovendien worden synthetische chemicaliën typisch gebruikt om kunstmatige reacties kunstmatig in te leidenOf pathogen virulentie. Hoewel de aanvulling van zulke chemische verbindingen die weerspiegelen van concentraties in planta mogelijk is, is deze toevoeging niet in de geringe mate van diffusie van wortel exudaten in de omringende rhizosfeer, die een chemotactische gradiënt genereert door microbes 28 , 32 genereert. Gezien de beperkingen van conventionele benaderingen om plant-microbe-interacties te bestuderen, kan de nauwkeurigheid en diepte van de verkregen gegevens worden belemmerd en restrictief, en de kennis die voortvloeit uit de conventionele benaderingen kan niet direct in planta vertaald worden. Veel aspecten van planten- Agrobacterium signalering zijn nog niet volledig begrepen, vooral in het vroege stadium van interacties, wanneer de ziektebeelden nog niet zijn ontwikkeld.
Om de beperkingen van conventionele benaderingen te wijzigen, presenteert dit werk een goedkope, goed beheersbare en flexibele hydroponische cOcultivatie systeem dat de onderzoekers in staat stelt diepere inzichten te krijgen in de complexe signalering- en reactiewegen bij de eerste fase van moleculaire plant-microbe interacties. Hydroponics is veel gebruikt om plantaardige voedingsstoffen, wortel exudaten, groeivoorwaarden en de effecten van metallische toxiciteit op planten 33 , 34 te bestuderen. Er zijn verschillende voordelen van hydroponische modellen, met inbegrip van de kleine ruimtelijke eisen, de toegankelijkheid van verschillende plantenweefsels, de strakke controle van de nutriënten / milieuomstandigheden en de bestrijding van de plaag / ziekte. Hydroponische systemen zijn ook minder beperkend voor plantengroei in vergelijking met agar / phytoagar plating technieken, die de groei meestal beperken na 2-3 weken. Belangrijk is dat het onderhoud van planten in de planten de natuurlijke wortesecretie vereiste die nodig is voor microbiële chemotaxis en virulentieinductie 8 , 29 . Het systeem beschrijftBed hier is eenvoudiger en minder arbeidsintensief dan de alternatieven 33 , 34 . Het maakt gebruik van minder onderdelen en vereist geen andere gereedschappen dan standaardscharen. Het gebruikt metaalnetwerk (in tegenstelling tot nylon 33 ) als een sterke ondersteuning voor plantengroei en een eenvoudige methode van beluchting onder steriele omstandigheden door schudden om microbiële groei te ondersteunen. Daarnaast kan het systeem metaalmaas van verschillende maten gebruiken om plantengroei te ondersteunen, die diverse plantensoorten herbergt zonder de breedte van hun wortels te beperken.
In het hier gepresenteerde hydroponische cocultivatiesysteem worden planten gekweekt in een steriel hydroponisch systeem waar de plantenwortels organische verbindingen scheiden die de groei van geïmpliceerde bacteriën ondersteunen. In dit cocultivatiesysteem worden geen kunstmatige chemicaliën, zoals plantaardige hormonen, verdedigingselectie, of virulentie-inducerende chemicaliën aangevuld, die de natuurlijke cel weerspiegeltSignalerende homeostase tijdens plant-microbe interacties. Met dit hydroponische cocultivatiesysteem was het mogelijk om gen expressie in Arabidopsis thaliana Col-0 wortelweefsel gelijktijdig te bepalen bij infectie door Agrobacterium , evenals de activatie van Agrobacterium genen bij cocultivatie met Arabidopsis . Verder werd aangetoond dat dit systeem geschikt is om Agrobacterium- bijlage aan plantenwortels te bestuderen, evenals het wortelgeheimprofiel van de plant, bij cocultivatie (infectie) met Agrobacterium ( Figuur 1 ).
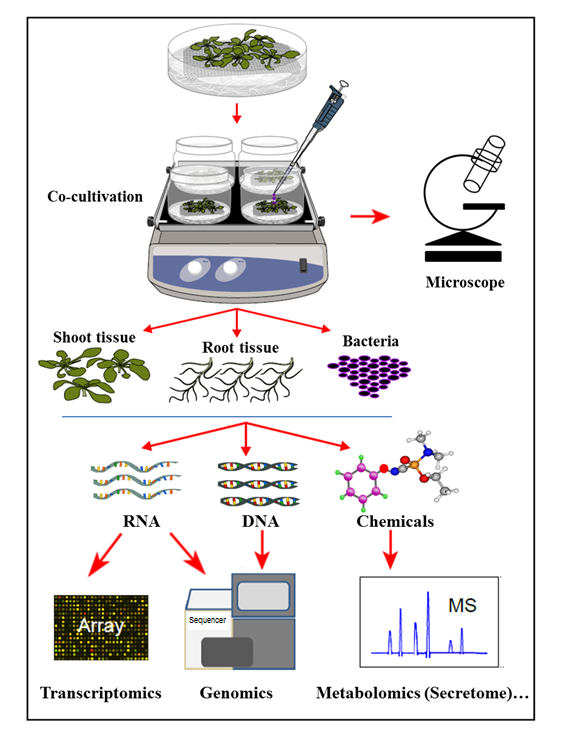
Figuur 1: Overzicht van het Hydroponic Cocultivation System, met voorbeeldanalyses. Planten worden op de maas gekweekt (scheuten boven het maas), met de wortels gedompeld in hydroponisch medium dat vervolgens met bacteriën fOf coculture. Plantweefsels en bacteriën worden vervolgens gescheiden voor gelijktijdige extracties en analyses. Dit cijfer is gewijzigd van referentie 35 .
